1203 Chapter 27. The Reproductive System
27.1 Anatomy and Physiology of the Male Reproductive System
Learning Objectives
By the end of this section, you will be able to:
- Describe the structure and function of the organs of the male reproductive system
- Describe the structure and function of the sperm cell
- Explain the events during spermatogenesis that produce haploid sperm from diploid cells
- Identify the importance of testosterone in male reproductive function

Unique for its role in human reproduction, a gamete is a specialized sex cell carrying 23 chromosomes—one half the number in body cells. At fertilization, the chromosomes in one male gamete, called a sperm (or spermatozoon), combine with the chromosomes in one female gamete, called an oocyte. The function of the male reproductive system (Figure 1) is to produce sperm and transfer them to the female reproductive tract. The paired testes are a crucial component in this process, as they produce both sperm and androgens, the hormones that support male reproductive physiology. In humans, the most important male androgen is testosterone. Several accessory organs and ducts aid the process of sperm maturation and transport the sperm and other seminal components to the penis, which delivers sperm to the female reproductive tract. In this section, we examine each of these different structures, and discuss the process of sperm production and transport.
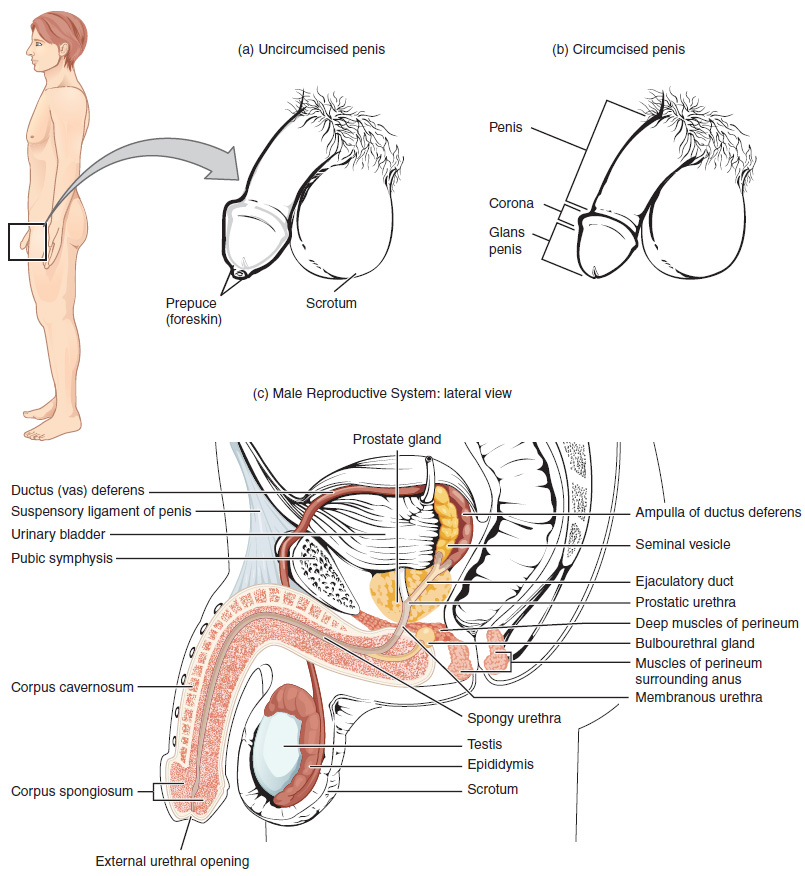
Scrotum
The testes are located in a skin-covered, highly pigmented, muscular sack called the scrotum that extends from the body behind the penis (see Figure 1). This location is important in sperm production, which occurs within the testes, and proceeds more efficiently when the testes are kept 2 to 4°C below core body temperature.
The dartos muscle makes up the subcutaneous muscle layer of the scrotum (Figure 2). It continues internally to make up the scrotal septum, a wall that divides the scrotum into two compartments, each housing one testis. Descending from the internal oblique muscle of the abdominal wall are the two cremaster muscles, which cover each testis like a muscular net. By contracting simultaneously, the dartos and cremaster muscles can elevate the testes in cold weather (or water), moving the testes closer to the body and decreasing the surface area of the scrotum to retain heat. Alternatively, as the environmental temperature increases, the scrotum relaxes, moving the testes farther from the body core and increasing scrotal surface area, which promotes heat loss. Externally, the scrotum has a raised medial thickening on the surface called the raphae.
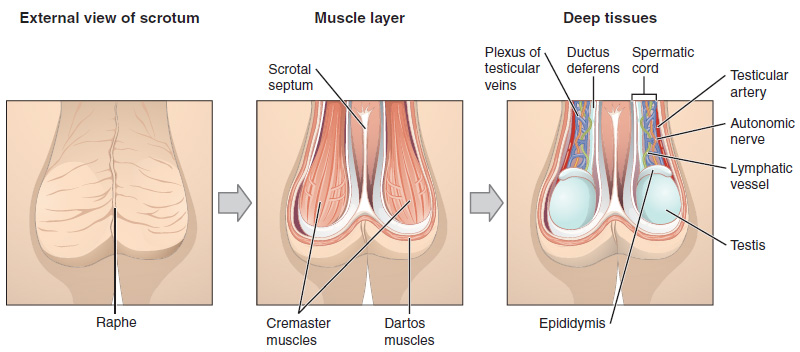
Testes
The testes (singular = testis) are the male gonads—that is, the male reproductive organs. They produce both sperm and androgens, such as testosterone, and are active throughout the reproductive lifespan of the male.
Paired ovals, the testes are each approximately 4 to 5 cm in length and are housed within the scrotum (see Figure 2). They are surrounded by two distinct layers of protective connective tissue (Figure 3). The outer tunica vaginalis is a serous membrane that has both a parietal and a thin visceral layer. Beneath the tunica vaginalis is the tunica albuginea, a tough, white, dense connective tissue layer covering the testis itself. Not only does the tunica albuginea cover the outside of the testis, it also invaginates to form septa that divide the testis into 300 to 400 structures called lobules. Within the lobules, sperm develop in structures called seminiferous tubules. During the seventh month of the developmental period of a male fetus, each testis moves through the abdominal musculature to descend into the scrotal cavity. This is called the “descent of the testis.” Cryptorchidism is the clinical term used when one or both of the testes fail to descend into the scrotum prior to birth.
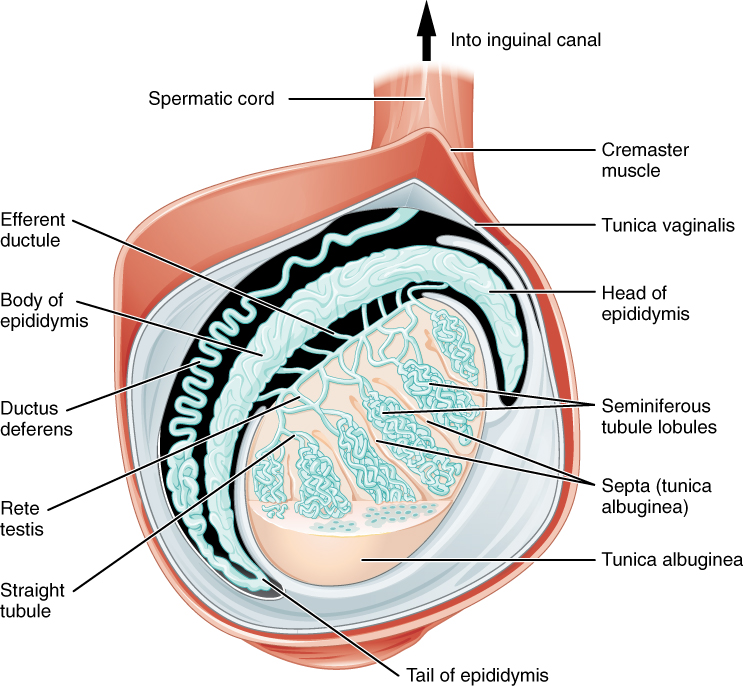
The tightly coiled seminiferous tubules form the bulk of each testis. They are composed of developing sperm cells surrounding a lumen, the hollow center of the tubule, where formed sperm are released into the duct system of the testis. Specifically, from the lumens of the seminiferous tubules, sperm move into the straight tubules (or tubuli recti), and from there into a fine meshwork of tubules called the rete testes. Sperm leave the rete testes, and the testis itself, through the 15 to 20 efferent ductules that cross the tunica albuginea.
Inside the seminiferous tubules are six different cell types. These include supporting cells called sustentacular cells, as well as five types of developing sperm cells called germ cells. Germ cell development progresses from the basement membrane—at the perimeter of the tubule—toward the lumen. Let’s look more closely at these cell types.
Sertoli Cells
Surrounding all stages of the developing sperm cells are elongate, branching Sertoli cells. Sertoli cells are a type of supporting cell called a sustentacular cell, or sustenocyte, that are typically found in epithelial tissue. Sertoli cells secrete signaling molecules that promote sperm production and can control whether germ cells live or die. They extend physically around the germ cells from the peripheral basement membrane of the seminiferous tubules to the lumen. Tight junctions between these sustentacular cells create the blood–testis barrier, which keeps bloodborne substances from reaching the germ cells and, at the same time, keeps surface antigens on developing germ cells from escaping into the bloodstream and prompting an autoimmune response.
Germ Cells
The least mature cells, the spermatogonia (singular = spermatogonium), line the basement membrane inside the tubule. Spermatogonia are the stem cells of the testis, which means that they are still able to differentiate into a variety of different cell types throughout adulthood. Spermatogonia divide to produce primary and secondary spermatocytes, then spermatids, which finally produce formed sperm. The process that begins with spermatogonia and concludes with the production of sperm is called spermatogenesis.
Spermatogenesis
As just noted, spermatogenesis occurs in the seminiferous tubules that form the bulk of each testis (see Figure 3). The process begins at puberty, after which time sperm are produced constantly throughout a man’s life. One production cycle, from spermatogonia through formed sperm, takes approximately 64 days. A new cycle starts approximately every 16 days, although this timing is not synchronous across the seminiferous tubules. Sperm counts—the total number of sperm a man produces—slowly decline after age 35, and some studies suggest that smoking can lower sperm counts irrespective of age.
The process of spermatogenesis begins with mitosis of the diploid spermatogonia (Figure 4). Because these cells are diploid (2n), they each have a complete copy of the father’s genetic material, or 46 chromosomes. However, mature gametes are haploid (1n), containing 23 chromosomes—meaning that daughter cells of spermatogonia must undergo a second cellular division through the process of meiosis.
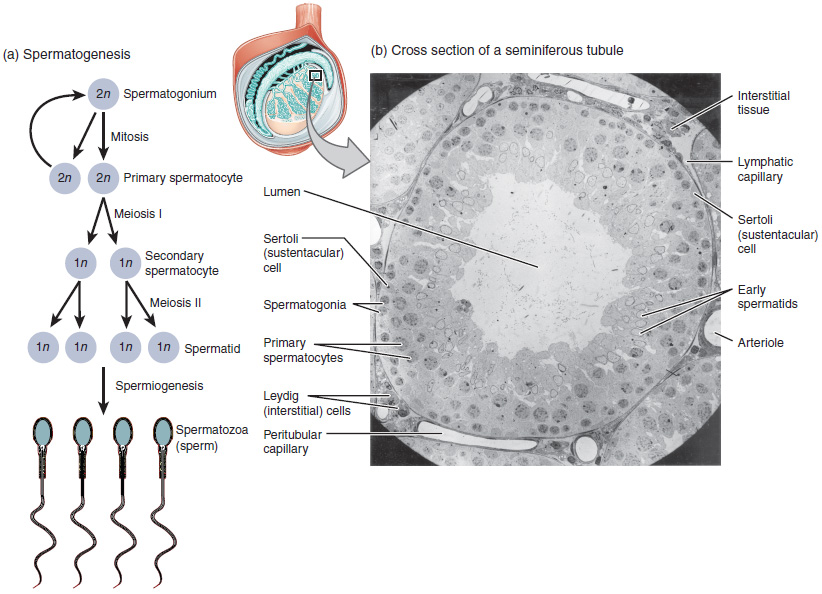
Two identical diploid cells result from spermatogonia mitosis. One of these cells remains a spermatogonium, and the other becomes a primary spermatocyte, the next stage in the process of spermatogenesis. As in mitosis, DNA is replicated in a primary spermatocyte, and the cell undergoes cell division to produce two cells with identical chromosomes. Each of these is a secondary spermatocyte. Now a second round of cell division occurs in both of the secondary spermatocytes, separating the chromosome pairs. This second meiotic division results in a total of four cells with only half of the number of chromosomes. Each of these new cells is a spermatid. Although haploid, early spermatids look very similar to cells in the earlier stages of spermatogenesis, with a round shape, central nucleus, and large amount of cytoplasm. A process called spermiogenesis transforms these early spermatids, reducing the cytoplasm, and beginning the formation of the parts of a true sperm. The fifth stage of germ cell formation—spermatozoa, or formed sperm—is the end result of this process, which occurs in the portion of the tubule nearest the lumen. Eventually, the sperm are released into the lumen and are moved along a series of ducts in the testis toward a structure called the epididymis for the next step of sperm maturation.
Structure of Formed Sperm
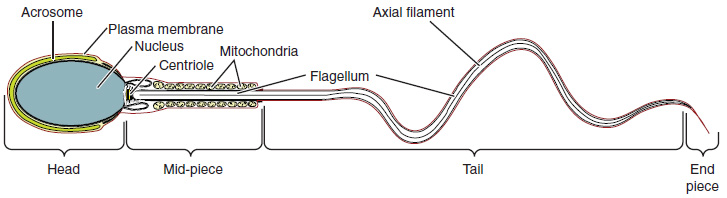
Sperm are smaller than most cells in the body; in fact, the volume of a sperm cell is 85,000 times less than that of the female gamete. Approximately 100 to 300 million sperm are produced each day, whereas women typically ovulate only one oocyte per month as is true for most cells in the body, the structure of sperm cells speaks to their function. Sperm have a distinctive head, mid-piece, and tail region (Figure 5). The head of the sperm contains the extremely compact haploid nucleus with very little cytoplasm. These qualities contribute to the overall small size of the sperm (the head is only 5 μm long). A structure called the acrosome covers most of the head of the sperm cell as a “cap” that is filled with lysosomal enzymes important for preparing sperm to participate in fertilization. Tightly packed mitochondria fill the mid-piece of the sperm. ATP produced by these mitochondria will power the flagellum, which extends from the neck and the mid-piece through the tail of the sperm, enabling it to move the entire sperm cell. The central strand of the flagellum, the axial filament, is formed from one centriole inside the maturing sperm cell during the final stages of spermatogenesis.
Sperm Transport
To fertilize an egg, sperm must be moved from the seminiferous tubules in the testes, through the epididymis, and—later during ejaculation—along the length of the penis and out into the female reproductive tract.
Role of the Epididymis
From the lumen of the seminiferous tubules, the immotile sperm are surrounded by testicular fluid and moved to the epididymis (plural = epididymides), a coiled tube attached to the testis where newly formed sperm continue to mature (see Figure 3). Though the epididymis does not take up much room in its tightly coiled state, it would be approximately 6 m (20 feet) long if straightened. It takes an average of 12 days for sperm to move through the coils of the epididymis, with the shortest recorded transit time in humans being one day. Sperm enter the head of the epididymis and are moved along predominantly by the contraction of smooth muscles lining the epididymal tubes. As they are moved along the length of the epididymis, the sperm further mature and acquire the ability to move under their own power. Once inside the female reproductive tract, they will use this ability to move independently toward the unfertilized egg. The more mature sperm are then stored in the tail of the epididymis (the final section) until ejaculation occurs.
Duct System
During ejaculation, sperm exit the tail of the epididymis and are pushed by smooth muscle contraction to the ductus deferens (also called the vas deferens). The ductus deferens is a thick, muscular tube that is bundled together inside the scrotum with connective tissue, blood vessels, and nerves into a structure called the spermatic cord (see Figure 1 and Figure 2). Because the ductus deferens is physically accessible within the scrotum, surgical sterilization to interrupt sperm delivery can be performed by cutting and sealing a small section of the ductus (vas) deferens. This procedure is called a vasectomy, and it is an effective form of male birth control. Although it may be possible to reverse a vasectomy, clinicians consider the procedure permanent, and advise men to undergo it only if they are certain they no longer wish to father children.

From each epididymis, each ductus deferens extends superiorly into the abdominal cavity through the inguinal canal in the abdominal wall. From here, the ductus deferens continues posteriorly to the pelvic cavity, ending posterior to the bladder where it dilates in a region called the ampulla (meaning “flask”).
Sperm make up only 5 percent of the final volume of semen, the thick, milky fluid that the male ejaculates. The bulk of semen is produced by three critical accessory glands of the male reproductive system: the seminal vesicles, the prostate, and the bulbourethral glands.
Seminal Vesicles
As sperm pass through the ampulla of the ductus deferens at ejaculation, they mix with fluid from the associated seminal vesicle (see Figure 1). The paired seminal vesicles are glands that contribute approximately 60 percent of the semen volume. Seminal vesicle fluid contains large amounts of fructose, which is used by the sperm mitochondria to generate ATP to allow movement through the female reproductive tract.
The fluid, now containing both sperm and seminal vesicle secretions, next moves into the associated ejaculatory duct, a short structure formed from the ampulla of the ductus deferens and the duct of the seminal vesicle. The paired ejaculatory ducts transport the seminal fluid into the next structure, the prostate gland.
Prostate Gland
As shown in Figure 1, the centrally located prostate gland sits anterior to the rectum at the base of the bladder surrounding the prostatic urethra (the portion of the urethra that runs within the prostate). About the size of a walnut, the prostate is formed of both muscular and glandular tissues. It excretes an alkaline, milky fluid to the passing seminal fluid—now called semen—that is critical to first coagulate and then decoagulate the semen following ejaculation. The temporary thickening of semen helps retain it within the female reproductive tract, providing time for sperm to utilize the fructose provided by seminal vesicle secretions. When the semen regains its fluid state, sperm can then pass farther into the female reproductive tract.
The prostate normally doubles in size during puberty. At approximately age 25, it gradually begins to enlarge again. This enlargement does not usually cause problems; however, abnormal growth of the prostate, or benign prostatic hyperplasia (BPH), can cause constriction of the urethra as it passes through the middle of the prostate gland, leading to a number of lower urinary tract symptoms, such as a frequent and intense urge to urinate, a weak stream, and a sensation that the bladder has not emptied completely. By age 60, approximately 40 percent of men have some degree of BPH. By age 80, the number of affected individuals has jumped to as many as 80 percent. Treatments for BPH attempt to relieve the pressure on the urethra so that urine can flow more normally. Mild to moderate symptoms are treated with medication, whereas severe enlargement of the prostate is treated by surgery in which a portion of the prostate tissue is removed.
Another common disorder involving the prostate is prostate cancer. According to the Centers for Disease Control and Prevention (CDC), prostate cancer is the second most common cancer in men. However, some forms of prostate cancer grow very slowly and thus may not ever require treatment. Aggressive forms of prostate cancer, in contrast, involve metastasis to vulnerable organs like the lungs and brain. There is no link between BPH and prostate cancer, but the symptoms are similar. Prostate cancer is detected by a medical history, a blood test, and a rectal exam that allows physicians to palpate the prostate and check for unusual masses. If a mass is detected, the cancer diagnosis is confirmed by biopsy of the cells.
Bulbourethral Glands
The final addition to semen is made by two bulbourethral glands (or Cowper’s glands) that release a thick, salty fluid that lubricates the end of the urethra and the vagina, and helps to clean urine residues from the penile urethra. The fluid from these accessory glands is released after the male becomes sexually aroused, and shortly before the release of the semen. It is therefore sometimes called pre-ejaculate. It is important to note that, in addition to the lubricating proteins, it is possible for bulbourethral fluid to pick up sperm already present in the urethra, and therefore it may be able to cause pregnancy.

The Penis
The penis is the male organ of copulation (sexual intercourse). It is flaccid for non-sexual actions, such as urination, and turgid and rod-like with sexual arousal. When erect, the stiffness of the organ allows it to penetrate into the vagina and deposit semen into the female reproductive tract.
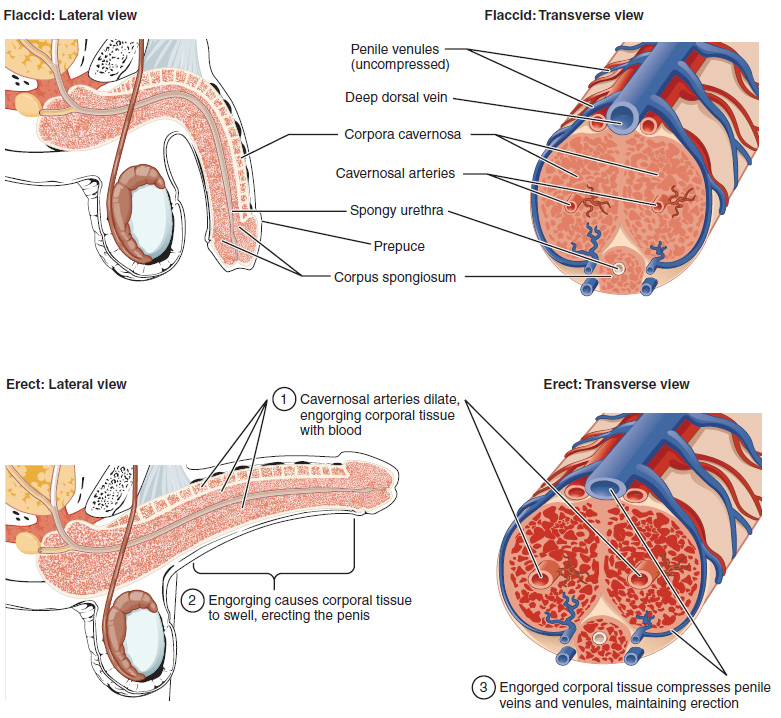
The shaft of the penis surrounds the urethra (Figure 6). The shaft is composed of three column-like chambers of erectile tissue that span the length of the shaft. Each of the two larger lateral chambers is called a corpus cavernosum (plural = corpora cavernosa). Together, these make up the bulk of the penis. The corpus spongiosum, which can be felt as a raised ridge on the erect penis, is a smaller chamber that surrounds the spongy, or penile, urethra. The end of the penis, called the glans penis, has a high concentration of nerve endings, resulting in very sensitive skin that influences the likelihood of ejaculation (see Figure 1). The skin from the shaft extends down over the glans and forms a collar called the prepuce (or foreskin). The foreskin also contains a dense concentration of nerve endings, and both lubricate and protect the sensitive skin of the glans penis. A surgical procedure called circumcision, often performed for religious or social reasons, removes the prepuce, typically within days of birth.
Both sexual arousal and REM sleep (during which dreaming occurs) can induce an erection. Penile erections are the result of vasocongestion, or engorgement of the tissues because of more arterial blood flowing into the penis than is leaving in the veins. During sexual arousal, nitric oxide (NO) is released from nerve endings near blood vessels within the corpora cavernosa and spongiosum. Release of NO activates a signaling pathway that results in relaxation of the smooth muscles that surround the penile arteries, causing them to dilate. This dilation increases the amount of blood that can enter the penis and induces the endothelial cells in the penile arterial walls to also secrete NO and perpetuate the vasodilation. The rapid increase in blood volume fills the erectile chambers, and the increased pressure of the filled chambers compresses the thin-walled penile venules, preventing venous drainage of the penis. The result of this increased blood flow to the penis and reduced blood return from the penis is erection. Depending on the flaccid dimensions of a penis, it can increase in size slightly or greatly during erection, with the average length of an erect penis measuring approximately 15 cm.
Male Reproductive SystemErectile dysfunction (ED) is a condition in which a man has difficulty either initiating or maintaining an erection. The combined prevalence of minimal, moderate, and complete ED is approximately 40 percent in men at age 40, and reaches nearly 70 percent by 70 years of age. In addition to aging, ED is associated with diabetes, vascular disease, psychiatric disorders, prostate disorders, the use of some drugs such as certain antidepressants, and problems with the testes resulting in low testosterone concentrations. These physical and emotional conditions can lead to interruptions in the vasodilation pathway and result in an inability to achieve an erection.
Recall that the release of NO induces relaxation of the smooth muscles that surround the penile arteries, leading to the vasodilation necessary to achieve an erection. To reverse the process of vasodilation, an enzyme called phosphodiesterase (PDE) degrades a key component of the NO signaling pathway called cGMP. There are several different forms of this enzyme, and PDE type 5 is the type of PDE found in the tissues of the penis. Scientists discovered that inhibiting PDE5 increases blood flow, and allows vasodilation of the penis to occur.
PDEs and the vasodilation signaling pathway are found in the vasculature in other parts of the body. In the 1990s, clinical trials of a PDE5 inhibitor called sildenafil were initiated to treat hypertension and angina pectoris (chest pain caused by poor blood flow through the heart). The trial showed that the drug was not effective at treating heart conditions, but many men experienced erection and priapism (erection lasting longer than 4 hours). Because of this, a clinical trial was started to investigate the ability of sildenafil to promote erections in men suffering from ED. In 1998, the FDA approved the drug, marketed as Viagra®. Since approval of the drug, sildenafil and similar PDE inhibitors now generate over a billion dollars a year in sales, and are reported to be effective in treating approximately 70 to 85 percent of cases of ED. Importantly, men with health problems—especially those with cardiac disease taking nitrates—should avoid Viagra or talk to their physician to find out if they are a candidate for the use of this drug, as deaths have been reported for at-risk users.
Testosterone
Testosterone, an androgen, is a steroid hormone produced by Leydig cells. The alternate term for Leydig cells, interstitial cells, reflects their location between the seminiferous tubules in the testes. In male embryos, testosterone is secreted by Leydig cells by the seventh week of development, with peak concentrations reached in the second trimester. This early release of testosterone results in the anatomical differentiation of the male sexual organs. In childhood, testosterone concentrations are low. They increase during puberty, activating characteristic physical changes and initiating spermatogenesis.
Functions of Testosterone
The continued presence of testosterone is necessary to keep the male reproductive system working properly, and Leydig cells produce approximately 6 to 7 mg of testosterone per day. Testicular steroidogenesis (the manufacture of androgens, including testosterone) results in testosterone concentrations that are 100 times higher in the testes than in the circulation. Maintaining these normal concentrations of testosterone promotes spermatogenesis, whereas low levels of testosterone can lead to infertility. In addition to intratesticular secretion, testosterone is also released into the systemic circulation and plays an important role in muscle development, bone growth, the development of secondary sex characteristics, and maintaining libido (sex drive) in both males and females. In females, the ovaries secrete small amounts of testosterone, although most is converted to estradiol. A small amount of testosterone is also secreted by the adrenal glands in both sexes.
Control of Testosterone
The regulation of testosterone concentrations throughout the body is critical for male reproductive function. The intricate interplay between the endocrine system and the reproductive system is shown in Figure 7.
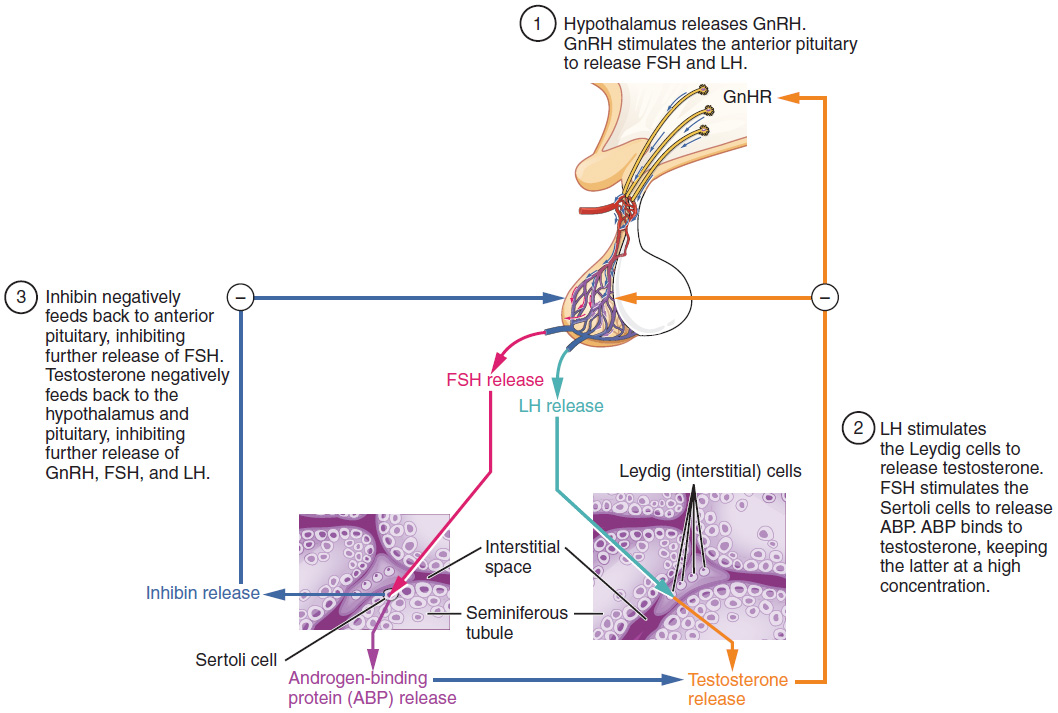
The regulation of Leydig cell production of testosterone begins outside of the testes. The hypothalamus and the pituitary gland in the brain integrate external and internal signals to control testosterone synthesis and secretion. The regulation begins in the hypothalamus. Pulsatile release of a hormone called gonadotropin-releasing hormone (GnRH) from the hypothalamus stimulates the endocrine release of hormones from the pituitary gland. Binding of GnRH to its receptors on the anterior pituitary gland stimulates release of the two gonadotropins: luteinizing hormone (LH) and follicle-stimulating hormone (FSH). These two hormones are critical for reproductive function in both men and women. In men, FSH binds predominantly to the Sertoli cells within the seminiferous tubules to promote spermatogenesis. FSH also stimulates the Sertoli cells to produce hormones called inhibins, which function to inhibit FSH release from the pituitary, thus reducing testosterone secretion. These polypeptide hormones correlate directly with Sertoli cell function and sperm number; inhibin B can be used as a marker of spermatogenic activity. In men, LH binds to receptors on Leydig cells in the testes and upregulates the production of testosterone.
A negative feedback loop predominantly controls the synthesis and secretion of both FSH and LH. Low blood concentrations of testosterone stimulate the hypothalamic release of GnRH. GnRH then stimulates the anterior pituitary to secrete LH into the bloodstream. In the testis, LH binds to LH receptors on Leydig cells and stimulates the release of testosterone. When concentrations of testosterone in the blood reach a critical threshold, testosterone itself will bind to androgen receptors on both the hypothalamus and the anterior pituitary, inhibiting the synthesis and secretion of GnRH and LH, respectively. When the blood concentrations of testosterone once again decline, testosterone no longer interacts with the receptors to the same degree and GnRH and LH are once again secreted, stimulating more testosterone production. This same process occurs with FSH and inhibin to control spermatogenesis.
