Unit 3.2: Viruses
Learning Objectives
After reading the following, you should be able to:
- Describe the characteristics of viruses.
- Explain why viruses are classified as obligatory intracellular parasites.
- Describe the general structure of an animal virion and a bacteriophage
- Describe characteristics that are used to classify viruses
- Describe the life cycle of the several types of animal viruses
- Describe how viruses can cause cancer.
- Contrast acute, latent and persistent viral infections and provide examples for each type.
Despite their small size, which prevented them from being seen with light microscopes, the discovery of a filterable component smaller than a bacterium that causes tobacco mosaic disease (TMD) dates back to 1892. At that time, Dmitri Ivanovski, a Russian botanist, discovered the source of TMD by using a porcelain filtering device designed to remove bacteria. An extract obtained from TMD-infected tobacco plants was made to determine the cause of the disease. Initially, the source of the disease was thought to be bacterial. It was surprising to everyone when Ivanovski, using a Chamberland filter, found that the cause of TMD was not removed after passing the extract through the porcelain filter. So if a bacterium was not the cause of TMD, what could be causing the disease? Ivanovski concluded the cause of TMD must be an extremely small bacterium or bacterial spore. Other scientists, including Martinus Beijerinck, continued investigating the cause of TMD. It was Beijerinck, in 1899, who eventually concluded the causative agent was not a bacterium but, instead, possibly a chemical, like a biological poison we would describe today as a toxin. As a result, the word virus, Latin for poison, was used to describe the cause of TMD a few years after Ivanovski’s initial discovery. Even though he was not able to see the virus that caused TMD, and did not realize the cause was not a bacterium, Ivanovski is credited as the original discoverer of viruses and a founder of the field of virology.
Today, we can see viruses using electron microscopes (Figure 3.13) and we know much more about them. Viruses are distinct biological entities; however, their evolutionary origin is still a matter of speculation. In terms of taxonomy, they are not included in the tree of life because they are acellular (not consisting of cells). In order to survive and reproduce, viruses must infect a cellular host, making them obligate intracellular parasites. The genome of a virus enters a host cell and directs the production of the viral components, proteins and nucleic acids, needed to form new virus particles called virions. New virions are made in the host cell by assembly of viral components. The new virions transport the viral genome to another host cell to carry out another round of infection. Table 3.2 summarizes the properties of viruses.

| Characteristics of Viruses |
|---|
| Infectious, acellular pathogens |
| Obligate intracellular parasites with host and cell-type specificity |
| DNA or RNA genome (never both) |
| Genome is surrounded by a protein capsid and, in some cases, a phospholipid memberane studded with viral glycoproteins |
| Lack genes for many products needed for successful reproduction, requiring exploitation of host-cell genomes to reproduce |
Viruses can infect every type of host cell, including those of plants, animals, fungi, protists, bacteria, and archaea. Most viruses will only be able to infect the cells of one or a few species of organism. This is called the host range. However, having a wide host range is not common and viruses will typically only infect specific hosts and only specific cell types within those hosts. The viruses that infect bacteria are called bacteriophages, or simply phages. The word phage comes from the Greek word for devour. Other viruses are just identified by their host group, such as animal or plant viruses. Once a cell is infected, the effects of the virus can vary depending on the type of virus. Viruses may cause abnormal growth of the cell or cell death, alter the cell’s genome, or cause little noticeable effect in the cell. In humans, a wide variety of viruses are capable of causing various infections and diseases. Some of the deadliest emerging pathogens in humans are viruses, yet we have few treatments or drugs to deal with viral infections, making them difficult to eradicate.
Viral Structures: In general, virions (viral particles) are small and cannot be observed using a regular light microscope. They are much smaller than prokaryotic and eukaryotic cells; this is an adaptation allowing viruses to infect these larger cells (see Figure 3.14). The size of a virion can range from 20 nm for small viruses up to 900 nm for typical, large viruses (see Figure 3.15). Recent discoveries, however, have identified new giant viral species, such as Pandoravirus salinus and Pithovirus sibericum, with sizes approaching that of a bacterial cell.

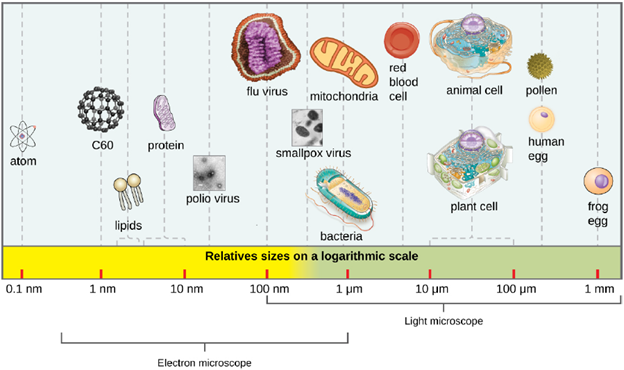
In 1935, after the development of the electron microscope, Wendell Stanley was the first scientist to crystallize the structure of the tobacco mosaic virus and discovered that it is composed of RNA and protein. In 1943, he isolated Influenza B virus, which contributed to the development of an influenza (flu) vaccine. Stanley’s discoveries unlocked the mystery of the nature of viruses that had been puzzling scientists for over 40 years and his contributions to the field of virology led to him being awarded the Nobel Prize in 1946.
As a result of continuing research into the nature of viruses, we now know they consist of a nucleic acid (either RNA or DNA, but never both) surrounded by a protein coat called a capsid (see Figure 3.16). The interior of the capsid is not filled with cytosol, as in a cell, but instead it contains the bare necessities in terms of genome and enzymes needed to direct the synthesis of new virions. Each capsid is composed of protein subunits called capsomeres made of one or more different types of capsomere proteins that interlock to form the closely packed capsid.
There are two categories of viruses based on general composition. Viruses formed from only a nucleic acid and capsid are called naked viruses or nonenveloped viruses. Viruses formed with a nucleic-acid packed capsid surrounded by a lipid layer are called enveloped viruses (see Figure 3.16). The viral envelope is a small portion of phospholipid membrane obtained as the virion buds from a host cell. The viral envelope may either be intracellular or cytoplasmic in origin.
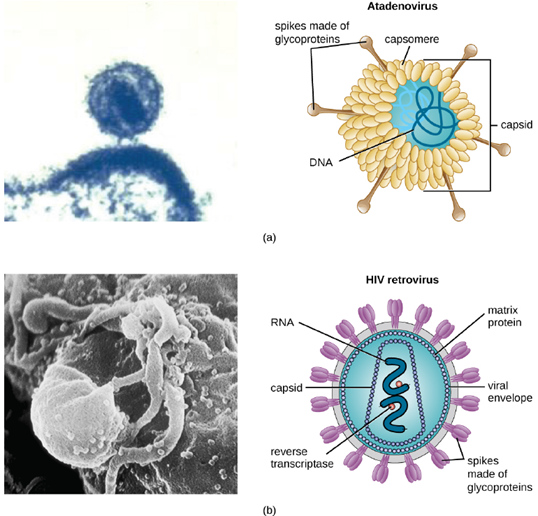
Extending outward and away from the capsid on some naked viruses and enveloped viruses are protein structures called spikes. At the tips of these spikes are structures that allow the virus to attach and enter a cell, like the influenza virus hemagglutinin spikes (H) or enzymes like the neuraminidase (N) influenza virus spikes that allow the virus to detach from the cell surface during release of new virions. Influenza viruses are often identified by their H and N spikes. For example, H1N1 influenza viruses were responsible for the pandemics in 1918 and 2009, H2N2 for the pandemic in 1957, and H3N2 for the pandemic in 1968.
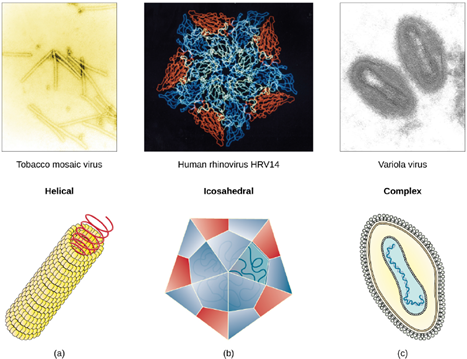
Viruses vary in the shape of their capsids, which can be either helical, polyhedral, or complex. A helical capsid forms the shape of tobacco mosaic virus (TMV), a naked helical virus, and Ebola virus, an enveloped helical virus. The capsid is cylindrical or rod shaped, with the genome fitting just inside the length of the capsid. Polyhedral capsids form the shapes of poliovirus and rhinovirus, and consist of a nucleic acid surrounded by a polyhedral (many sided) capsid in the form of an icosahedron. An icosahedral capsid is a three-dimensional, 20-sided structure with 12 vertices. These capsids somewhat resemble a soccer ball. Both helical and polyhedral viruses can have envelopes. Viral shapes seen in certain types of bacteriophages, such as T4 phage, and poxviruses, like vaccinia virus, may have features of both polyhedral and helical viruses so they are described as a complex viral shape (see Figure 3.17). In the bacteriophage complex form, the genome is located within the polyhedral head and the sheath connects the head to the tail fibers and tail pins that help the virus attach to receptors on the host cell’s surface. Poxviruses that have complex shapes are often brick shaped, with intricate surface characteristics not seen in the other categories of capsid.
Classification and Taxonomy of Viruses: Although viruses are not classified in the three domains of life, their numbers are great enough to require classification. Since 1971, the International Union of Microbiological Societies Virology Division has given the task of developing, refining, and maintaining universal virus taxonomy to the International Committee on Taxonomy of Viruses (ICTV). Since viruses can mutate so quickly, it can be difficult to classify them into a genus and a species epithet using the binomial nomenclature system. Thus, the ICTV’s viral nomenclature system classifies viruses into families and genera based on viral genetics, chemistry, morphology, and mechanism of multiplication. To date, the ICTV has classified known viruses in seven orders, 96 families, and 350 genera. Viral family names end in –viridae (e.g, Parvoviridae) and genus names end in −virus (e.g., Parvovirus). The names of viral orders, families, and genera are all italicized. When referring to a viral species, we often use a genus and species epithet such as Pandoravirus dulcis or Pandoravirus salinus. Often times, we just use the common name for the virus, which is not italicized.
Aside from formal systems of nomenclature, viruses are often informally grouped into categories based on chemistry, morphology, or other characteristics they share in common. Categories may include naked or enveloped structure, single-stranded (ss) or double-stranded (ds) DNA or ss or ds RNA genomes, segmented or non-segmented genomes, and positive-strand (+) or negative-strand (−) RNA. For example, herpes viruses can be classified as a dsDNA enveloped virus; human immunodeficiency virus (HIV) is a +ssRNA enveloped virus, and tobacco mosaic virus is a +ssRNA virus. Other characteristics such as host specificity, tissue specificity, capsid shape, and special genes or enzymes may also be used to describe groups of similar viruses.
The Viral Lifecycle: All viruses depend on cells for reproduction and metabolic processes. By themselves, viruses do not encode for all of the enzymes necessary for viral replication. But within a host cell, a virus can commandeer cellular machinery to produce more viral particles. In eukaryotic cells, most DNA viruses can replicate inside the nucleus, with an exception observed in the large DNA viruses, such as the poxviruses, that can replicate in the cytoplasm. RNA viruses that infect animal cells often replicate in the cytoplasm.
Life Cycle of Viruses with Animal Hosts: Attachment is the first stage in the infection process in which the animal virus interacts with specific surface receptors found on the target host cells.
After binding to host receptors, animal viruses enter through cell-mediated endocytosis (engulfment by the host cell) or through membrane fusion (viral envelope with the host cell membrane). Many viruses are host specific, meaning they only infect a certain type of host; and most viruses only infect certain types of cells within tissues. This specificity is called a tissue tropism. Examples of this are demonstrated by the poliovirus, which exhibits tropism for the tissues of the brain and spinal cord, or the influenza virus, which has a primary tropism for the respiratory tract.
As the capsids of animal viruses enter the host cell, either via cell-mediated endocytosis or fusion, the capsid must be removed to release the viral nucleic acids. This process is known as uncoating. Separation of the nucleic acid from the protein coat can occur through different mechanisms depending on the virus. Some examples include: The capsid being digested by host enzymes, the capsid being digested by viral enzymes and the capsid dissociating due to the internal pH of the cell. This step does not occur in bacteriophages as they inject their nucleic acid into their host cell, leaving the capsid outside.
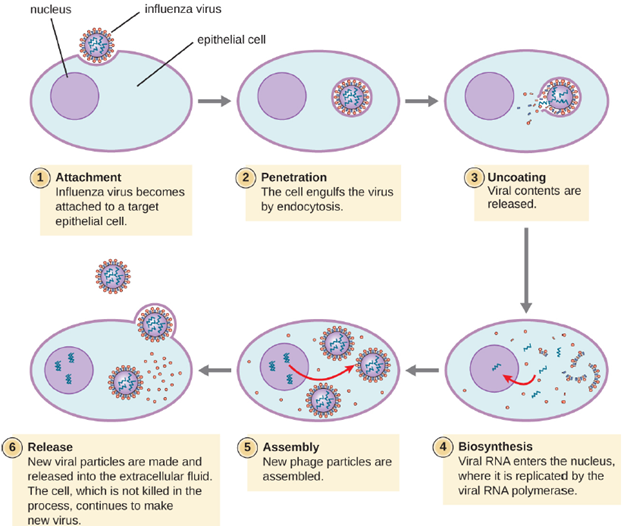
Animal viruses do not always express their genes using the normal flow of genetic information—from DNA to RNA to protein. Some viruses have a dsDNA genome like cellular organisms and can follow the normal flow. However, others may have ssDNA, dsRNA, or ssRNA genomes. The nature of the genome determines how the genome is replicated and expressed as viral proteins. If a genome is ssDNA, host enzymes will be used to synthesize a second strand that is complementary to the genome strand, thus producing dsDNA. The dsDNA can now be replicated, transcribed, and translated similar to host DNA.
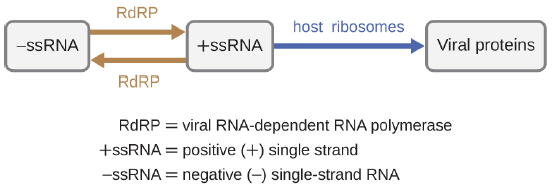
If the viral genome is RNA, a different mechanism must be used. There are three types of RNA genome: dsRNA, positive (+) single-strand (+ssRNA) or negative (−) single-strand RNA (−ssRNA). If a virus has a +ssRNA genome, it can be translated directly to make viral proteins. Viral genomic +ssRNA acts like cellular mRNA. However, if a virus contains a −ssRNA genome, the host ribosomes cannot translate it until the −ssRNA is replicated into +ssRNA by viral RNA-dependent RNA polymerase (RdRP) (see Figure 3.19). The RdRP is brought in by the virus and can be used to make +ssRNA from the original −ssRNA genome. The RdRP is also an important enzyme for the replication of dsRNA viruses, because it uses the negative strand of the double-stranded genome as a template to create +ssRNA. The newly synthesized +ssRNA copies can then be translated by cellular ribosomes.
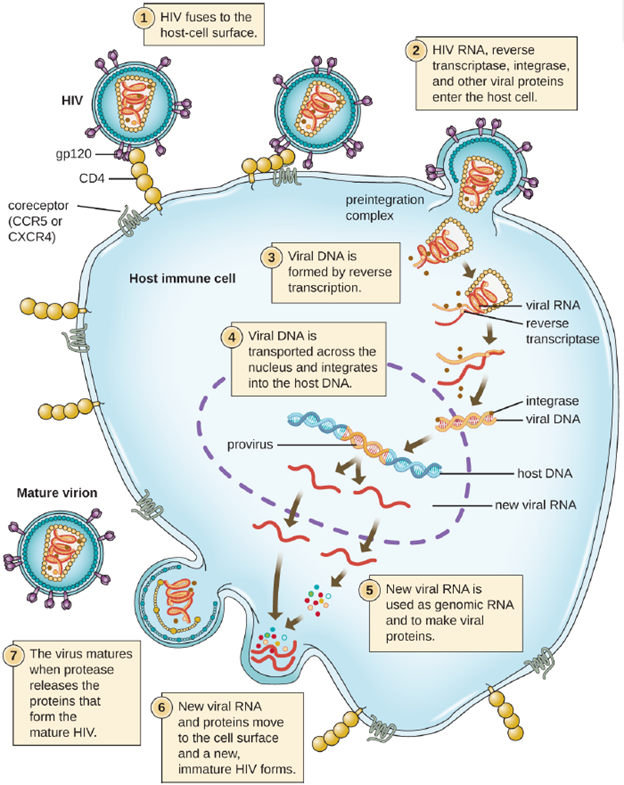
An alternative mechanism for viral nucleic acid synthesis is observed in the retroviruses, which are +ssRNA viruses (see Figure 3.20). Single-stranded RNA viruses such as HIV carry a special enzyme called reverse transcriptase within the capsid that synthesizes a complementary ssDNA (cDNA) copy using the +ssRNA genome as a template. The ssDNA is then made into dsDNA, which can integrate into the host chromosome and become a permanent part of the host. The integrated viral genome is called a provirus. The virus now can remain in the host for a long time to establish a chronic infection. The provirus stage is similar to the prophage stage in a bacterial infection during the lysogenic cycle. However, unlike prophage, the provirus does not undergo excision after splicing into the genome.
Regardless of the mechanism of biosynthesis, once the viral components have been produced they undergo maturation and self-assemble into new virions. These new virions must be released from the cell so they can infect new hosts. While some viruses, mostly those that are non-enveloped, can be released by triggering cell lysis, as is the case with bacteriophages, animal viruses also can leave the cell by budding, which is used by enveloped virus, and exocytosis, which is used by non-enveloped virus. In budding, the virion takes a part of the cells membrane as it leaves, using it to form its viral membrane. In exocytosis, the new virion is excreted from the cell in a vesicle that merges with the plasma membrane releasing the virion outside the cell. In both of these methods of release, the cell does not die and can continue to produce new virions over months or even years.
Persistent Infections: Most viruses cause acute infections, whereby the virus is rapidly produced causing symptoms over a brief period of time, but other modes of infection exist. Persistent infection occurs when a virus is not completely cleared from the system of the host but stays in certain tissues or organs of the infected person. The virus may remain silent or undergo productive infection without seriously harming or killing the host. Mechanisms of persistent infection may involve the regulation of the viral or host gene expressions or the alteration of the host immune response. The two primary categories of persistent infections are latent infection and chronic infection. Examples of viruses that cause latent infections include herpes simplex virus (oral and genital herpes), varicella-zoster virus (chickenpox and shingles), and Epstein-Barr virus (mononucleosis). Hepatitis C virus and HIV are two examples of viruses that cause long-term chronic infections.
- Latent Infection: Not all animal viruses undergo replication by the lytic cycle. There are viruses that are capable of remaining hidden or dormant inside the cell in a process called latency. These types of viruses are known as latent viruses and may cause latent infections. Viruses capable of latency may initially cause an acute infection before becoming dormant. Latent viruses may remain dormant by existing as circular viral genome molecules outside of the host chromosome. Others become proviruses by integrating into the host genome. During dormancy, viruses do not cause any symptoms of disease and may be difficult to detect. A patient may be unaware that he or she is carrying the virus unless a viral diagnostic test has been performed.
- Chronic Infection: A chronic infection is a disease with symptoms that are recurrent or persistent over a long time. Some viral infections can be chronic if the body is unable to eliminate the virus. HIV is an example of a virus that produces a chronic infection, often after a long period of latency. Once a person becomes infected with HIV, the virus can be detected in tissues continuously thereafter, but untreated patients often experience no symptoms for years. However, the virus maintains chronic persistence through several mechanisms that interfere with immune function, including preventing expression of viral antigens on the surface of infected cells, altering immune cells themselves, restricting expression of viral genes, and rapidly changing viral antigens through mutation. Eventually, the damage to the immune system results in progression of the disease leading to acquired immunodeficiency syndrome (AIDS). The various mechanisms that HIV uses to avoid being cleared by the immune system are also used by other chronically infecting viruses, including the hepatitis C virus.
Viruses and Cancer: Viruses can cause cancer by transforming a normal cell into a malignant cell. A direct oncogenic (cancer causing) viral mechanism involves either the insertion of additional viral oncogenic genes into the host cell, or the enhancement of already existing oncogenic genes in the genome. Tumor viruses come in a variety of forms. Viruses with a DNA genome, such as adenovirus, and viruses with an RNA genome, like the hepatitis C virus (HCV), can cause cancers. Retroviruses having both DNA and RNA genomes (human T-lymphotropic virus and hepatitis B virus) can also cause cancers. Viruses can also become carcinogenic when they integrate into the host cell genome as part of a biological accident, such as polyomaviruses and papillomaviruses. Worldwide, cancer viruses are estimated to cause 15-20% of all cancers in humans. Most viral infections, however, do not lead to tumor formation; several factors influence the progression from viral infection to cancer development. These factors include host’s genetic makeup, mutation occurrence, exposure to cancer causing agents, and immune impairment.
Viruses typically initiate cancer development by suppressing the host’s immune system, causing inflammation over a long period of time, or by altering host genes. Cancer cells have characteristics that differ from normal cells, such as acquiring the ability to grow uncontrollably. This can result from having control of their own growth signals, losing sensitivity to anti-growth signals, and losing the ability to undergo apoptosis, or programmed cell death. Cancer cells don’t experience biological aging, and maintain their ability to undergo cell division and growth. Transformation occurs when a virus infects and genetically alters a cell. The infected cell is regulated by the viral genes and has the ability to undergo abnormal new growth. Scientists have been able to discern some commonality among viruses that cause tumors. The tumor viruses or oncoviruses change cells by integrating their genetic material with the host cell’s DNA. Unlike the integration seen in prophages, this is a permanent insertion; the genetic material is never removed. The insertion mechanism can differ depending on whether the nucleic acid in the virus is DNA or RNA. In DNA viruses, the genetic material can be directly inserted into the host’s DNA. RNA viruses must first transcribe RNA to DNA and then insert the genetic material into the host cell’s DNA.
