Main Body
Non-Mendelian Inheritance
So far in this course, we have seen that we can predict how traits are passed from parents to offspring using Mendel’s Laws of Inheritance (Self test: See if you can define all three, then refresh your memory with the glossary below) Unfortunately, in nature things are seldom so simple, and there are many cases where patterns of inheritance do not follow Mendel’s laws. In this chapter, we will take a look at several examples of non-Mendelian inheritance.
Mendel’s Laws of Inheritance
Non-Mendelian Dominance
In Mendel’s peas, every trait Mendel described exhibited complete dominance, meaning one allele would always mask the expression of the other. Since Mendel’s time, other types of Dominant/Recessive relationships have been described, including Co-Dominance and Incomplete Dominance.
Incomplete Dominance
To fully understand non-Mendelian modes of dominance, we need to first understand why alleles give rise to their associated traits. For example, we know that in Mendel’s peas, flower colour is controlled by a single gene with two alleles. One allele (Dominant) produces purple flowers, while the other (Recessive) produces white flowers. Why is this?
It turns out that this gene codes for a protein that is needed to produce anthocyanin pigment. The recessive allele codes for a non-functioning protein, and thus homozygous-recessive peas produce flowers with no anthocyanin pigment at all, giving them their white colour. If a plant has even one functioning copy of the allele, it can produce enough anthocyanin to completely dye its flowers purple, which is why heterozygous pea plants are still purple. This mechanism was described fairly recently, in 2010 (150 years after Mendel first described the phenotype!) If you want to learn more about the specifics of how this gene works, check out this article from New Zealand’s Science Learning Hub.
In this case, one functioning allele produces enough pigment to produce the purple phenotype. What if this wasn’t the case? What if a homozygous plant with two copies of the purple allele produced twice as much pigment as a heterozygous plant with only one copy? We will consider this scenario in the following exercise:
This scenario, where the phenotype that arises from a given set of alleles is dose-dependent, is an example of incomplete dominance, where the recessive allele is not completely masked by the dominant allele. This typically leads to a third phenotype in heterozygotes, which looks like a muted version of the homozygous-dominant phenotype.
Co-dominance
Co-dominance occurs when two alleles of the same gene are fully expressed at the same time in a heterozygous organism. This leads to a heterogeneous phenotype, where both parental phenotypes are expressed simultaneously and equally. This is distinct from incomplete dominance, where the phenotype of one allele only partially masks the phenotype of the other.
Hypothetical Case Study: Thorny Roses
Let’s imagine you are a geneticist in charge of breeding roses. The variety of rose you work with has two types of thorns, long (TL) and short (TS):
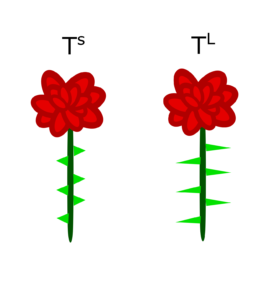
Luckily, you are able to acquire pure-breeding lines for each phenotype. You cross these lines to produce F1 offspring, all of which have the following phenotype:

Note that these F1 offspring both have short and long thorns in equal proportion. You then cross two F1 offspring and note the following phenotype ratios in the F2 offspring:
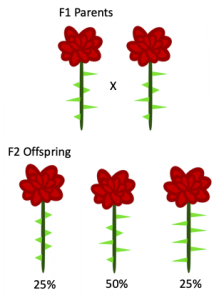
These offspring ratios tell you a few things about the thorn-size phenotype. Test your understanding using the following exercise:
Contrast this scenario with the non-Mendelian pea flowers we discussed above. Heterozygote pea flowers express an intermediate phenotype that resembles a blend of the parental phenotypes. In this scenario, heterozygote roses express both parental phenotypes simultaneously and equally. Thus, we can say that the alleles for thorn-size are co-dominant to each other.
Allelic Series
In all the scenarios we have considered so far, genes always have two alleles. In nature, it is common for genes to have more than two alleles, each with their own associated phenotype. In these cases, we say the gene exhibits multiple allelism.
Hypothetical Case Study: Blue-spot Butterflies
Imagine you are a geneticist studying the rare blue-spot butterfly (sadly not a real insect). This butterfly has three distinct wing phenotypes: Small, medium, and large.

Through previous testing, you know that this phenotype is controlled at a single locus. At first glance, this looks like it could be a classic case of incomplete dominance, where heterozygous individuals produce a “blended” phenotype. However, while rearing the butterflies in the lab, you realize you can establish pure-breeding lines of each phenotype! What does this tell you about the system?
Now that you have this new-found knowledge about the wing-size phenotype, you run the following test-crosses on pure-breeding lines of each phenotype:
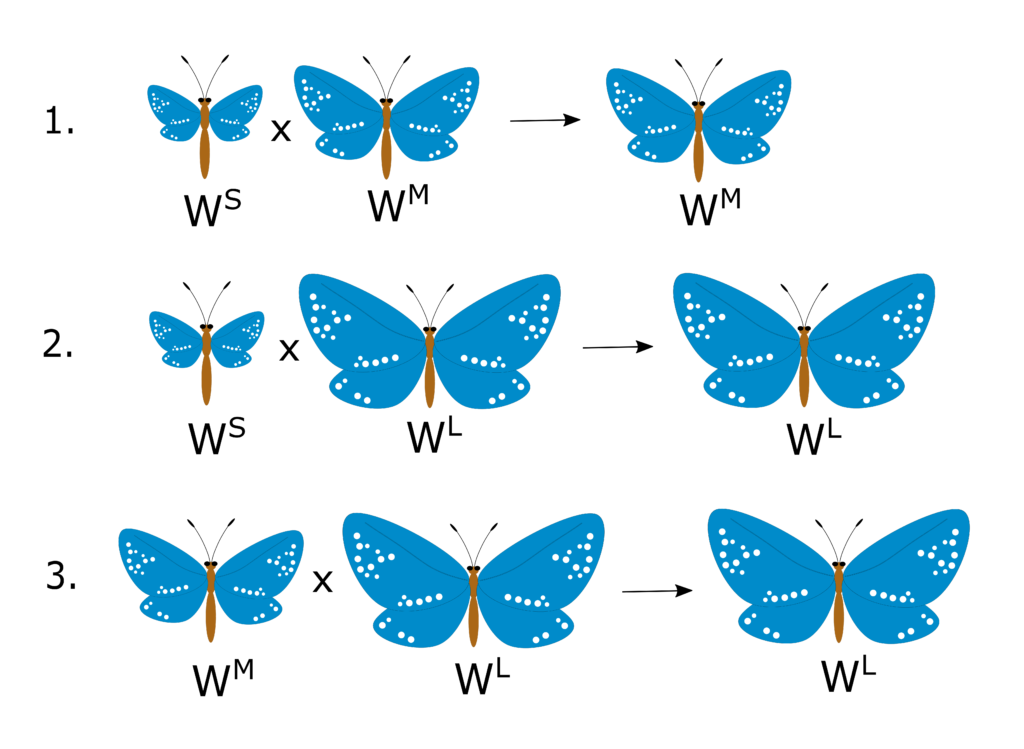
Now this is an odd pattern! Let’s try to analyse it one piece of information at a time:
- The first test cross tells us that WM is dominant over WS
- The second test cross tells us that WL is dominant over WS
- The third test cross tells us that WL is also dominant over WM
It appears that there is a hierarchy of dominance between the three alleles, structured as WL > WM > WS. What you’ve described here is an allelic series, a special case of multiple allelism where there is a set hierarchy for all the alleles involved.
Summary: Non-Mendelian Dominance
Here are brief definitions of the three modes of non-Mendelian dominance discussed above
- Incomplete dominance occurs when one copy of a dominant allele isn’t enough to completely mask the recessive phenotype.
- Co-dominance occurs when both alleles of a gene are expressed equally in a heterozygote.
- Allelic series occurs when there are more than two alleles for a given gene, and those alleles follow a set hierarchy of dominance.
These cases can sometimes be difficult to tell apart, particularly incomplete dominance and co-dominance. Remember that all phenotypes have a molecular basis: genes code for proteins, and it is the action of these proteins that lead to visible phenotypes. Often, we need to investigate both the genetic basis and the molecular basis for a phenotype before we can fully understand why certain patterns of inheritance arise.
Lethal Alleles
Some genes carry out functions that are absolutely essential to the survival of an organism. For example, genes that control the development of heart muscles, or self-recognition by the immune system. These are broadly termed Essential Genes, because loss of function in these genes would be lethal under normal circumstances. Change-of-function mutations in these genes are, naturally, referred to as Lethal Mutations, and alleles that carry lethal mutations are termed Lethal Alleles. It may seem counter-intuitive that lethal alleles could persist within a population, but they do appear from time to time, and they tend to come with their own set of non-Mendelian quirks.
Dominant Lethal Alleles
Dominant lethal alleles display complete dominance over the wild-type, making them difficult to study – even one copy leads to death. In most cases, these alleles aren’t passed from parent to offspring, but instead arise spontaneously from mutations. One exception to this is Huntington’s Disease:
Case Study: Huntington’s Disease
A common example of a dominant lethal allele in humans is Huntington’s Disease (HD). HD is a late-onset neuro-degenerative disease; the life expectancy for people with HD is typically 40-50 years. HD is caused by mutations in the gene Huntingtin (htt), and even one copy of the mutant allele is enough to cause full progression of the disease.
The late-onset of this disease is important: it allows a mutant copy of Huntingtin to be passed to offspring before the disease shows any sign of progression. Keep this example in mind; we’ll be coming back to it later in the module.
Recessive Lethal Alleles
Case Study: Cuénot’s yellow mice
You are working with a population of lab-reared mice. These mice can either have yellow or black fur, and you want to figure out how the gene for fur colour is inherited. While performing test crosses between yellow mice, you notice the following pattern emerge repeatedly in their offspring:
2 yellow : 1 black
You might deduce that the yellow phenotype is dominant over black, but where is this odd ratio coming from? Shouldn’t you be seeing 3:1 yellow to black mice? By performing more test crosses, you realize that it is impossible to establish pure-breeding lines of yellow mice, meaning it is impossible for mice to be homozygous for the fur colour gene.
This is the exact scenario encountered by the French geneticist Lucien Cuénot, who described this relationship in 1905. At the time, he didn’t know what caused this odd phenomenon, and it wasn’t until a few years later that his colleagues were able to tease it apart. It turns out that the gene for coat colour also controls a process that is essential to the development of mice embryos. The yellow allele of this gene carries a recessive mutation that alters mouse development, and kills the embryo. Thus, homozygous-yellow mice die in utero and can never be observed in test crosses!
For a more comprehensive look at this story, and references to the original papers, see this article from Nature.
In modern genetics terms, we would say that the allele for yellow fur is pleiotropic, meaning it codes for multiple phenotypes. In this case, the primary phenotype (coat colour) is dominant, while the secondary phenotype (death) is recessive.
This case is particularly interesting: The pleiotropy of the yellow allele is actually very important for the discovery of the recessive lethal phenotype; if the lethal allele wasn’t attached to a visible phenotype, we might never had noticed anything was wrong, aside from maybe reduced fecundity in our monohybrid crosses. In fact, most recessive lethal alleles aren’t connected to a dominant phenotype, and are completely invisible. Thus, heterozygotes can unknowingly distribute them throughout a population with no indication that they ever carry a mutation.
Conditional Lethality
This last case is a bit odd, and can be difficult to identify. Conditional Lethality occurs when a given allele produces a lethal phenotype, but that phenotype is only expressed under certain environmental conditions.
Case Study: Favism
Favism is a hereditary condition that affects certain people of Middle Eastern and Mediterranean descent. The condition is caused by a deficiency in the enzyme glucose-6-phosphate dehydrogenase, which is needed to breakdown sugars. This condition is dominant, X-linked, and is completely invisible most of the time.
If an individual with the condition eats certain foods (fava beans, specifically) or takes certain medications, they experience a massive breakdown of red blood cells. The effects of this range from mild (anemia) to severe (blood clots, liver failure), and in extreme cases can be lethal. On the other hand, this condition is also protective against malaria, which is why it has persisted in affected populations despite it’s disastrous side effects.
Conditional Expression of Alleles
As we just saw in the case of conditional lethality, phenotypes can sometimes be affected by environmental factors, independent of the genotype. This is actually very common in nature, and is broadly termed Conditional Expression of Alleles. Many organs and tissues rely on conditional expression of alleles for normal function; for example, cells in your stomach lining should only produces acid when there is food present that needs digesting. Otherwise, there is a risk of the acid burning through the mucous that protects your stomach lining.
Variable Expressivity and Penetrance
In all the cases we have seen so far – in both Mendelian and non-Mendelian modes of inheritance – individuals with a given genotype will always express the same phenotype. This, unfortunately, is not always the case.
Variable Expessivity
Expressivity tells us the degree to which a phenotype will be expressed for an individual with a particular genotype. In all the examples we have seen so far, all alleles have had 100% expressivity, meaning a specific genotype will produce a specific phenotype 100% of the time.
Alleles with Variable Expressivity will display a gradient of phenotypes within a population, with different individuals showing greater or lesser degrees of expression of the associated phenotype.
Variable Penetrance
Penetrance tells us the probability that a phenotype will be expressed given the presence of the associated genotype. Again, in all the examples we have seen so far, alleles have had 100% penetrance. Alleles with Variable Penetrance won’t always express their phenotypes, and so can’t be tracked using typical patterns of inheritance.

Figure 1: This figure shows examples of what variable expressivity and penetrance might look like in three different populations of red flowers. In box A, we see a typical Mendelian scenario: all the flowers have the exact same phenotype. In box B, we see an alternative scenario where the allele for red colouration has variable expressivity. In box C, the allele for red colouration has normal expressivity but variable penetrance. Remember, within a single box of this Figure, all the flowers have the exact same genotype.
Sex-Influenced Traits
You should already be familiar with sex-linked genes, which are found exclusively on sex chromosomes, and thus can show different inheritance patterns for males vs. females. However, not all traits that vary between sexes are caused by sex-linked genes: some are produced by autosomal genes! How can this be?
Sex-influenced traits arise because the expression of the phenotype requires the presence of specific sex hormones and\or products of sex-linked genes. For example, certain risk-alleles for breast cancer are autosomal, but are only activated in the presence of estrogen. Since men typically produce less estrogen, they are much less likely to exhibit the breast cancer phenotype than females. Other sex-influenced traits can show sex-related differences in which allele is dominant (e.g. for male-pattern baldness), or the timing or degree of phenotypic expression.
In some cases, the expression of an autosomal trait is never observed in one sex: we call these sex-limited traits, since the expression of the gene is limited to the other sex.
Mendel’s Laws are the product of an exceptional model system
Mendel was actually exceptionally lucky in choosing his experimental traits:
- All 7 of the traits he described in his pea plants are controlled by single genes,
- None of these genes are linked on the same chromosome,
- None of them are sex-linked,
- They all have exactly 2 alleles,
- They all exhibit complete dominance, complete penetrance, and complete expressivity.
As we saw in this chapter, these “regular” genetic traits do a very poor job at representing the true diversity of genetic traits in nature, and if we are going to study genes we need to understand and appreciate all the exceptions to these rules.
Alright that’s a lot of new material to chew on! Let’s see if we can synthesize it all together in the following exercises:
Exercise 1: Red-eared sliders
Exercise 2: Huntington’s Disease
Exercise 3: Red and White Petals
Anthocyanins are a family of organic pigments that are commonly found in plants. They range in colour from red to blue to purple.