Lecture Materials
Introduction to Entomology
Introduction
Entomology is the scientific study of insects. The word “entomon” (Greek) and the word “insectum”(Latin) both mean “divided”. The insect body is divided into recognizable parts; however, insects come in a startling diversity of forms. View this slideshow [http://www.alexanderwild.com/] to experience a tiny portion of insect diversity.
Diversity
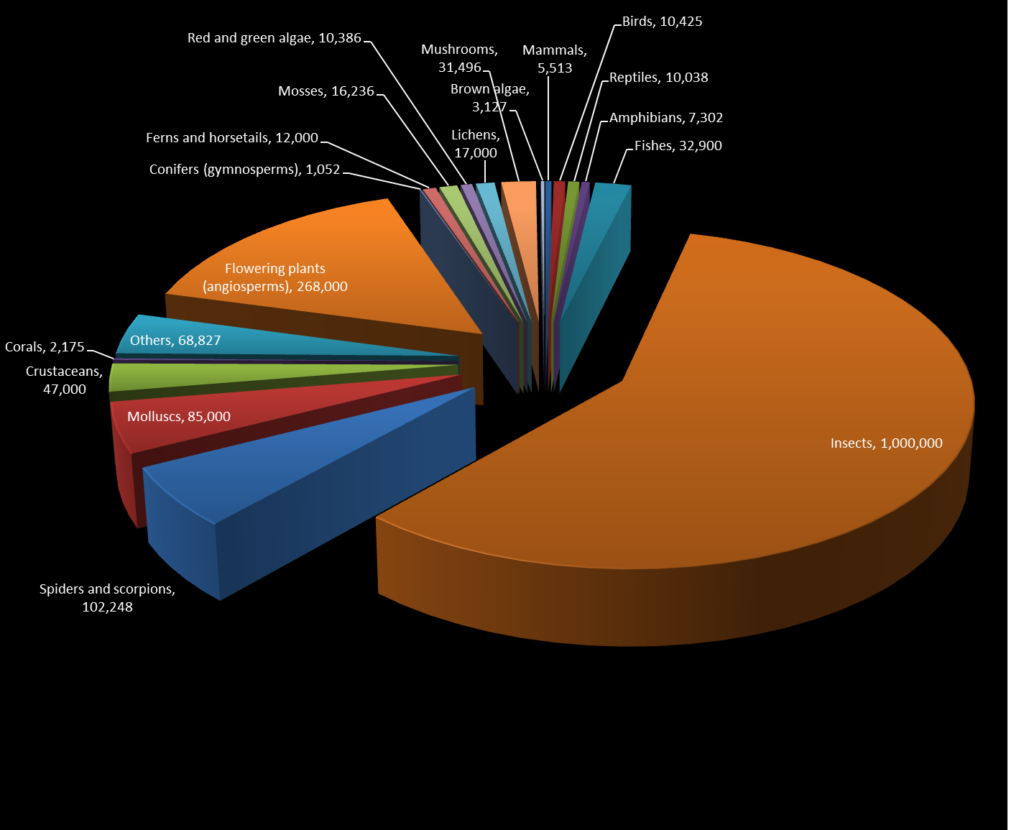
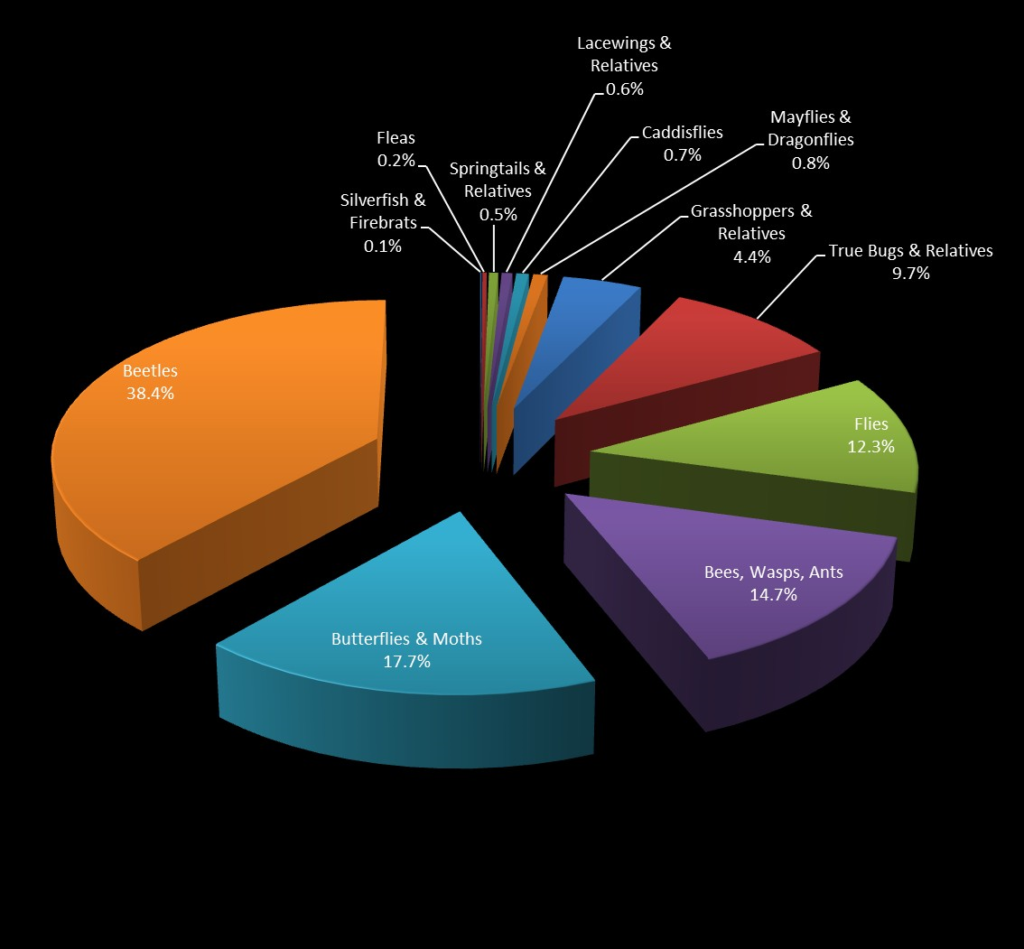
More than 1.7 million species of organisms have been described. About 1 million of those are insects (Fig. 1). The largest order of insects is the beetles, with over 390,000 species known. New species of organisms (including insects) are being found all the time, and it is likely that we have only described a fraction of the extant diversity: estimates of the number of species on Earth range from 5 million to 30 million or more.
Classification
With so many different species, we need a logical, systematic way to group similar organisms together. Better yet, we need a system that reflects evolutionary relationships, rather than simply physical similarities. The branch of biology that deals with taxonomy and nomenclature is called systematics. Taxonomy is the science of grouping organisms with a similar evolutionary history together, while nomenclature is the selection of names for those groups.
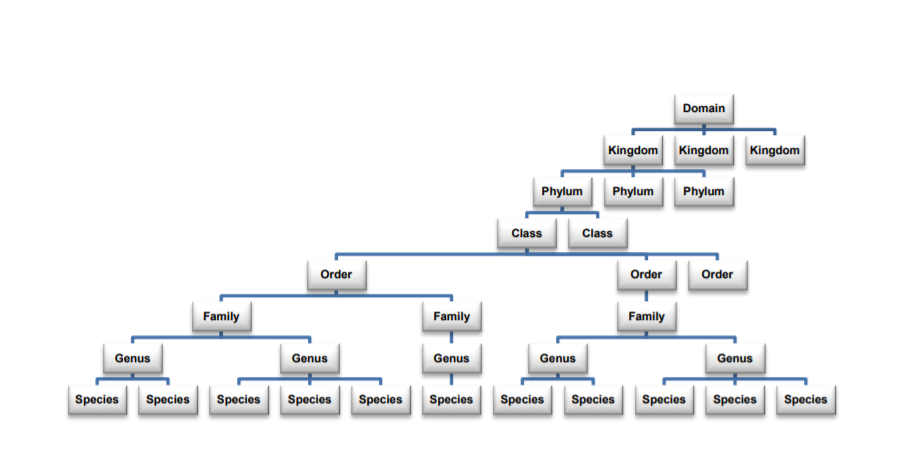
The system we use is based on a nested hierarchy devised by Carl von Linné in 1758. Smaller, more specific categories are nested within larger groupings. The highest modern level of this system is the Domain, and the most specific level is, of course, the species (Fig. 3).
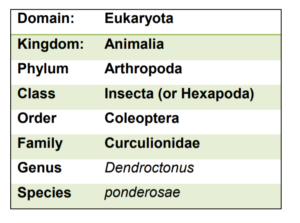
As an example, Table 1 shows the classification of one of the insects you will learn about in this course, the mountain pine beetle. Note that higher level group names, like Coleoptera, are always capitalized. A particular species is denoted by a binomial, made up of both the genus name and the species name. The genus name is always capitalized, while the species name is in lower case. Both the genus and species names are italicized, or underlined in handwriting.
Table 1. Classification of the mountain pine beetle, Dendroctonus ponderosae
Why do you have to learn the genus and species names instead of common names? For one thing, many organisms do not have common names. For another, common names can vary widely from place to place, and person to person, and they can be misleading. In a field like forest health, it is crucial to be sure you are dealing with a particular species and not a similar species (perhaps with a different host, biology, or life cycle) in order to choose the best management strategy and tactics.
Body Plan
Despite their enormous diversity, all insects share some common features. Like all arthropods, they have an exoskeleton and jointed legs. Insects are distinguished from other arthropods by having three distinct body regions called tagmata, each specialized for different functions (Fig. 4). The head is specialized for feeding, and for sensing the surrounding world. All the mouthparts are here, as well as one pair of antennae, and (often) a pair of large compound eyes. There may also be simple eyes, called ocelli, that detect light levels but are likely not image-forming. The thorax is specialized for locomotion, and bears three pairs of jointed legs and usually two pairs of wings (in adults). The abdomen is specialized for vital functions such as digestion and reproduction.
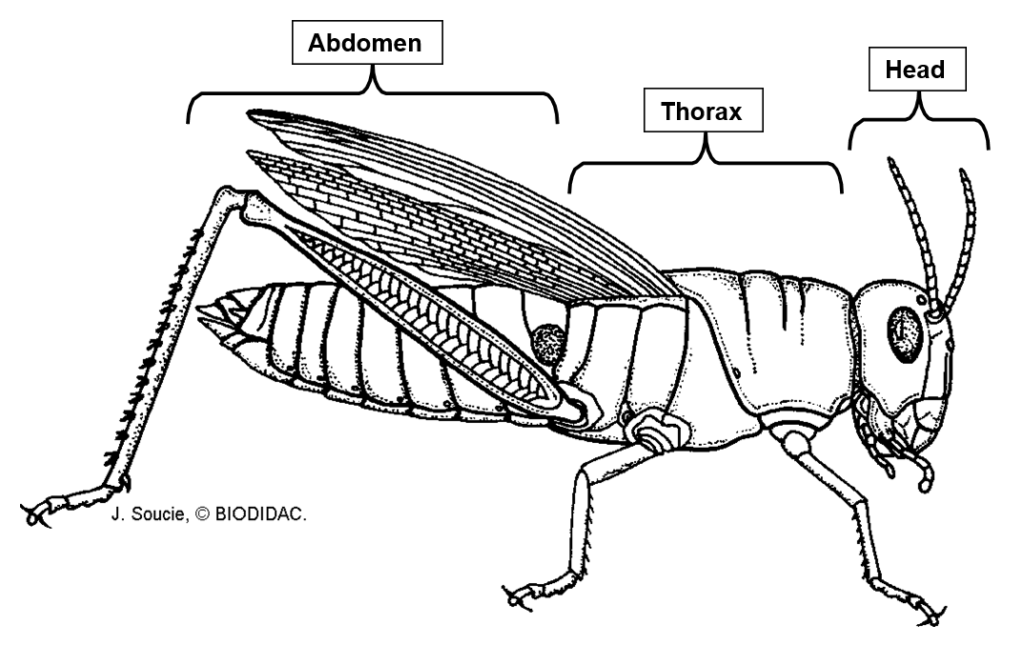
Exoskeleton
One of the keys to insect success is their exoskeleton. The skeleton provides support, protection, and sites for muscle attachment; unlike our skeleton though, the insect skeleton is on the outside surface of the body. It can form a tremendous variety of external shapes, while leaving the internal organs virtually unchanged. This is one of the reasons behind the diversity of insects that you saw earlier.
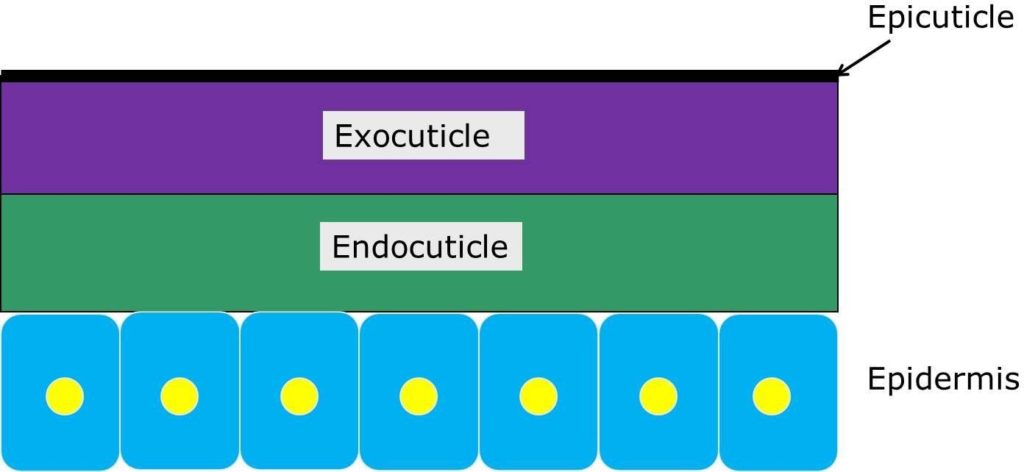
The exoskeleton, or cuticle, sits on top of the cellular epidermis, and is secreted by the epidermal cells (Fig. 5). The inner, flexible layer of cuticle is called the endocuticle. The outer layer, which is often rigid, is called the exocuticle. On top of the exocuticle is a thin, waxy layer called the epicuticle, which helps to protect the insect from desiccation and physical damage.
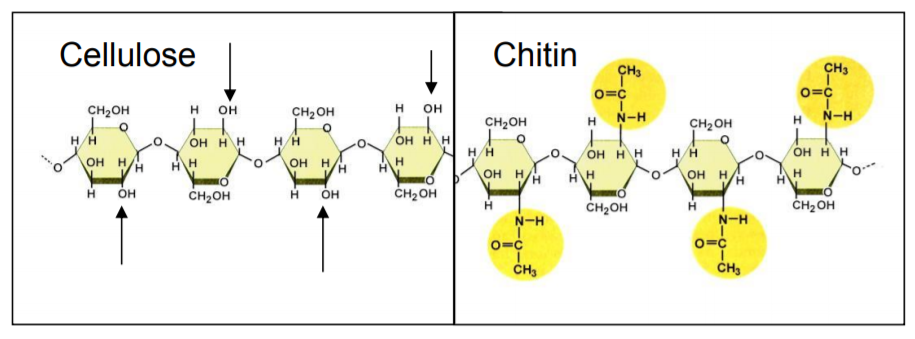
The insect exoskeleton is mostly made up of chitin, which is chemically similar to cellulose (Fig. 6). In areas where the exocuticle is rigid, the chitin has undergone sclerotization, or “tanning”, a process in which the chitin chains are cross-linked with a protein called sclerotin.
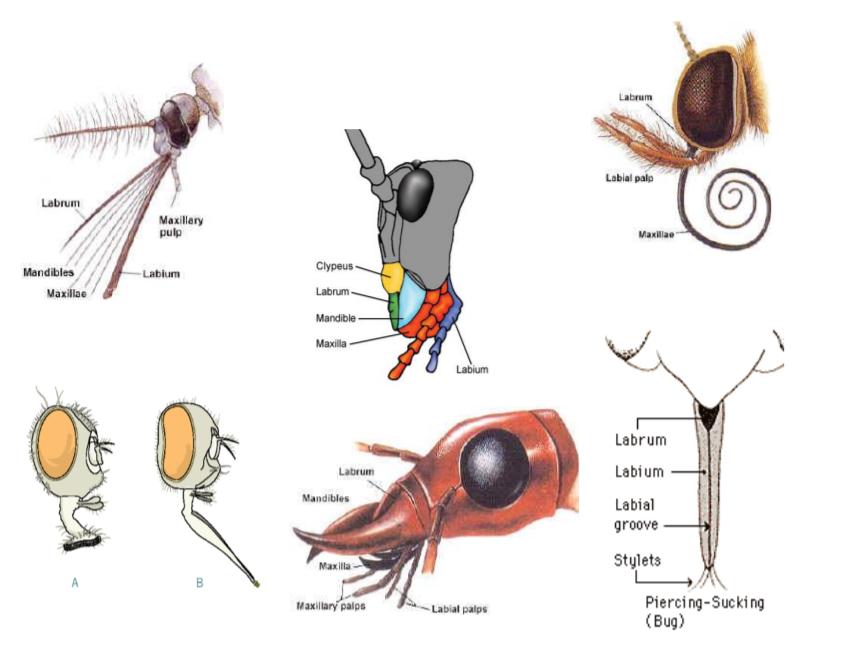
Mouthparts
Insects rely on a tremendous variety of food sources, and utilize many feeding modes. Different insect groups have mouthparts that are adapted for different types of foods (Fig. 7). If you are in FSTY 317, you will have an opportunity to take a closer look at some of these adaptations in the lab.
Legs
Insect legs are also adapted for life in many different environments (Fig. 8). As with mouthparts, even in leg types that look very different, all of the same parts are there but in modified shapes. Once again, if you are in FSTY 317, you will have an opportunity to take a closer look at some of these adaptations in the lab.
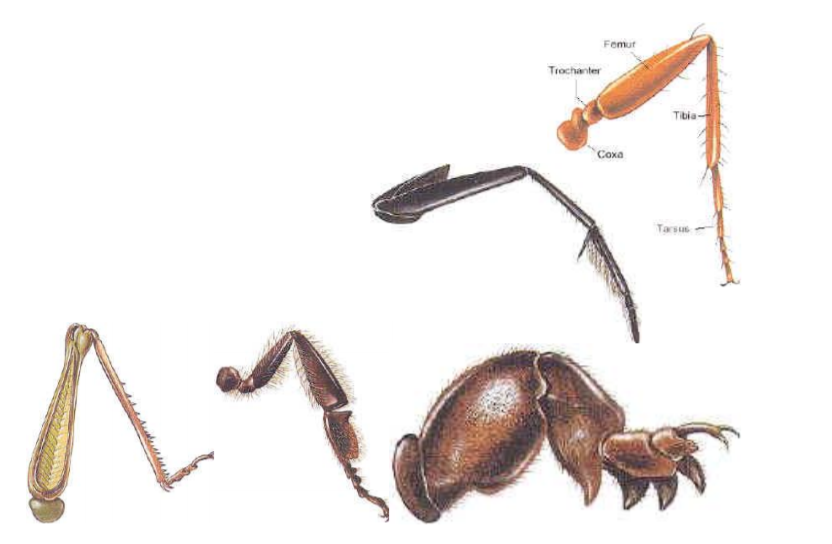
Wings
Perhaps the most important factor in the success and diversity of insects is their ability to fly. In fact almost all orders of insects end in “-ptera”, meaning “wing”. The names in some way describe the shape or structure of the wings: for example, the name of the order Coleoptera, or beetles, means “sheath wing”. Most insects, like this beetle https://upload.wikimedia.org/wikipedia/commons/7/7a/Lady_beetle_taking_flight_right_bright.jpg have two pairs of wings. Beetles have the front wings modified into hardened coverings for the membranous hind wings. These hardened front wings are called elytra (singular: elytron), the “sheath” referred to by the order name Coleoptera.
{kind=link}
Some well-known insects, such as monarch butterflies and some dragonflies, are capable of undertaking long migrations. Other species can also disperse long distances when they are carried by air currents and storm systems. Such systems carried mountain pine beetles from British Columbia, across the Rocky Mountains, and into Alberta.
Reproduction
Most insects reproduce sexually, although sex determination is not always the same as in humans. In some groups, such as the Lepidoptera (butterflies and moths), females have X and Y chromosomes, while males have two X chromosomes. Honey bees and some other social insects are haplodiploid. The females are diploid, carrying two complete sets of chromosomes, but males are haploid, and carry only one set. This system leads to a very high degree of relatedness among the members of a colony, all of whom are offspring of a single mated pair.
Many insects also reproduce asexually. Some, like several species of stick insects (https://upload.wikimedia.org/wikipedia/commons/e/ea/2013-03- 09_Indische_Stabschrecke_carausius_morosus_anagoria.JPG), are completely parthenogenic and may have been so for thousands or even millions of years. Others, like many aphids (Fig. 9), are parthenogenic for much of the summer, but produce males and reproduce sexually in the fall as environmental conditions start to deteriorate.
{kind=link}

Development
Insects have a tough, protective exoskeleton. In order to grow, they must moult, or shed, that exoskeleton and produce a new one (https://www.youtube.com/watch?v=wpNehJ0V_aM). In almost all insects, moulting happens only in the immature stages. The period of time between moults is called an instar. From the time the insect hatches from the egg until its first moult, it is in its first instar. From the first moult to the second moult is the second instar, and so on. The final moult transforms the insect to the adult stage.
There are three basic types of development patterns, or metamorphosis, in insects. The most ancestral insects change little from one moult to the next, except in size. At the final moult, they become reproductively mature, but they are always wingless. This type of metamorphosis is called ametabolous.
Other insects change relatively gradually. With each successive moult the wings become more developed; at the final moult, the wings are fully functional and the insect is reproductively mature. This type of gradual metamorphosis is called hemimetabolous (in insects with aquatic immature stages and terrestrial adult stages, such as dragonflies), and paurometabolous in terrestrial insects.
About 80% of insects are holometabolous, and undergo “complete” metamorphosis. The egg hatches into a larva, which will grow through several moults. At the second to last moult, it will become a pupa. In this stage, almost all of its tissues and cells are broken down and rebuilt into the adult insect. Holometabolous insects have adults that look completely different from their larvae, and that often inhabit completely different environments. In these insects, the larvae are specialized for feeding and growth, while the adults are specialized for dispersal and reproduction.
Remember that insects are small, and mostly ectothermic. Their rate of development, as well as their mobility and metabolic efficiency, are strongly influenced by the temperature of their surroundings.
Life Cycles
Some insects have relatively straightforward life cycles, while others are very complex (Fig. 10). Managing populations of forest health agents requires you to understand the life cycle of the species you want to manage. What is its reproductive biology, how many life stages are there, when and where does each life stage occur, how does each stage make its living? The answers to questions such as these determine what management strategies and tactics are likely to be most effective.
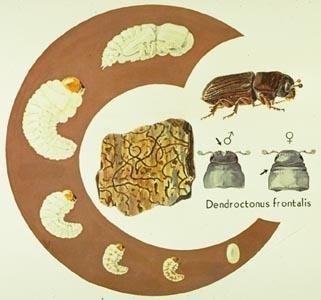
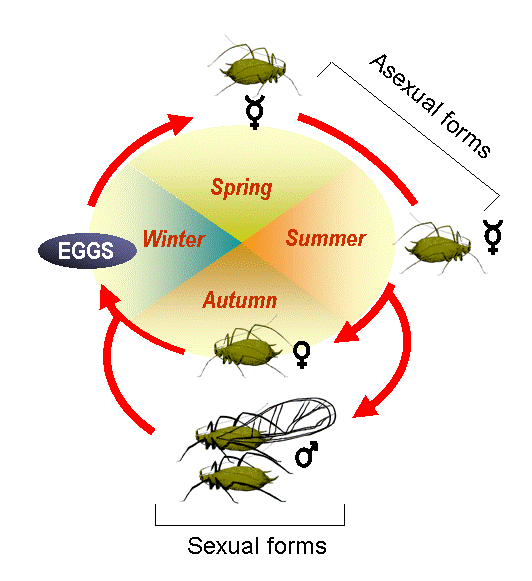
Factors Affecting Insect Populations
Populations of some insects are strongly regulated by the availability of susceptible hosts. Many bark beetles require dead, dying or downed trees of a particular species. As the availability of such host material increases, so does the bark beetle population. For example, spruce beetle (Dendroctonus rufipennis, http://www.forestpests.org/vd/images/0805006-SMPT.jpg) populations tend to increase following blow-down events, which result in large amounts of downed spruce in a stand. This type of population regulation is known as bottom-up regulation, and is commonly seen among K-selected organisms.
{kind=link}
Other insect populations are mostly regulated by factors other than food, such as parasites and predators. For example, Douglas-fir tussock moth (Orgyia pseudtsugata, https://tidcf.nrcan.gc.ca/images_web/imfc/insectes/moyen/orgia_pseudotsugata_21.jpg) larvae feed on the foliage of Douglas-fir. Food is rarely limiting, as Douglas-fir is common in the areas where this insect is found, and the trees produce copious amounts of foliage. However, as larval densities increase, so do the densities of parasites and predators that feed on the larvae, and various diseases are transmitted more easily in dense populations. Douglas-fir tussock moth populations usually decline sharply after one to three years because of the influence of parasites, predators and pathogens. This type of population regulation is known as top-down regulation, and is commonly seen among r-selected organisms.
{kind=link}
Feeding Groups
Given their diversity, it shouldn’t be surprising that insects can be found almost everywhere and can eat virtually anything. Many species, though, are restricted to certain types of food, certain species of food, or even certain parts of their food species. Most of the insects you will be concerned with in this course are phytophagous, or herbivorous, meaning that they eat plant material. Some of these are specialists on foliage (foliophagous), phloem (phloeophagous), or on wood (xylophagous).
Other insects feed primarily on fungi (mycophagous), or on dead material (saprophagous). Still others are predaceous, or predatory. Some are parasites of vertebrates or plants, keeping their hosts alive while they feed on them. Finally, there are some insects that adopt a strategy intermediate between parasitism and predation. These insects lay their eggs in or on other insects, and the larvae parasitize their hosts, keeping the host alive. At some point in development, however, the parasitic larvae switch to a predatory strategy, kill their host, and consume it.
Insect-Plant Interactions
On any plant (any tree, in this course), herbivores will be found feeding on it. Even pollinators of a flowering plant are included here, as they are feeding on nectar and pollen produced by the plant. There are also likely to be insects that are predators or parasitoids of the herbivores. A third category of insects is often referred to as “tourists”. They use trees for shelter, basking, or perches, but do not derive nutrients from the trees (directly or indirectly).
All plants have a wide range of physical and chemical defenses against herbivores. Physical defenses may range from droopy leaves, to trichomes, to various thorns and spines. Chemical defenses may make the plant smell like it is not susceptible, or may even act as repellents, anti-feedants or digestion inhibitors. They may also be outright toxic to herbivorous insects.
Forests and Insects
As hosts for insects, trees are very long-lived. A tree’s generation time far outlasts that of its insect herbivores. Furthermore, like other plants, trees can’t run away! They are dependent on physical and chemical defenses to reduce the level of damage caused by herbivores to the point where it does not threaten their survival or reproductive ability.
Herbivores are a natural part of a tree’s ecosystem. In general, trees have evolved to produce excess photosynthetic organs (leaves) to tolerate herbivore feeding, bark to prevent entry of organisms into the wood, and formidable chemical defenses throughout their systems. Some level of damage is likely to be of little or no consequence to the tree, unless it has other issues as well. For example, a tree that has been infected by a pathogen, or scarred by a fire, may be much more susceptible to bark beetles than trees that have not experienced these pre-disposing disturbances.
Ecological Effects
Within a forest, herbivorous insects play several important roles (Fig. 11). They are particularly important in nutrient cycling, breaking down plant tissues that can tie up nutrients for a long time. They may do this directly: for example, termites have symbiotic protozoans in their guts that allow them to digest the cellulose in wood: They may also release nutrients indirectly by feeding on foliage, then depositing scraps or feces on the forest floor.
As you have already seen elsewhere in the course, forest insects are important in successional processes. Even if they are acting on a small scale, the disturbances they cause lead to a mosaic of tree species and ages on the landscape.
Finally, forest insect activities help to provide food and habitat for many other organisms, including bears, woodpeckers, other insects (especially ants), and fungi. Dead trees are important components of forest systems.

Detection and Diagnosis
Forest insects are always present, in any ecosystem. Detecting their presence does NOT mean that they constitute a forest health problem. It is important that forest managers make objective responses based on sound diagnostic and ecological information. They must also always consider the management objectives and values for each particular stand. Insects, including those that feed on trees, are a natural part of the ecosystem, so you must learn to take a holistic view.
When diagnosing a potential forest insect pest problem, you will often observe symptoms. Theserepresent the tree’s response to a herbivorous insect or other disturbance agent. Generally, you will not be able to determine the exact cause of the symptoms, although they may help you to narrow down the potential causative agents. To diagnose the problem definitively, you will need to observe signs, which show direct evidence of the causative agent. Often, the best signs are the agents themselves. For example, a symptom might be a dying lodgepole pine in a young plantation in central BC. That symptom suggests that the root system or the stem has been damaged in some way, but it does not tell you exactly what caused the damage. If you dig around in the area of the root collar and find masses of resin-soaked soil with white grubs inside them, you can diagnose the damage as being caused by Warren root collar weevil, Hylobius warreni. Notice that factors such as location and tree species can also be important in diagnosis.
Hazard and Risk
When deciding whether or not to take action to manage a forest health agent, the two concepts of hazard and risk are important to consider. Unfortunately, the two terms are easy to confuse, and are frequently used interchangeably by many people.
The stand hazard generally refers to the susceptibility of a stand to damage. In other words, if a particular insect was to start causing damage in that stand, how bad would the damage be? Factors such as the location of the stand, the species mix, the age profile, and overall tree health can be important in determining stand hazard. Many of the major forest insect pests in BC have well-developed hazard rating systems that allow inputs of various data, and produce a hazard index for a given pest in a particular stand.
The stand risk generally refers to how likely an outbreak is to occur in that particular stand, often based on its proximity to the nearest large population of the pest of interest. Analyses of both hazard and risk are necessary to determine what management strategies and tactics might be most useful, or even whether management is indicated at all. Both hazard and risk, as I’ve described them, are often rolled into a single risk-rating system.
Hazard and risk are most useful as tools to evaluate future damage to a stand. In order to apply them successfully, managers must understand the ecology of the tree, the insect, the stand and the landscape. Both the tree’s requirements and the insect’s requirements will have an impact on future damage, and insect population dynamics can determine how likely an outbreak may be, and how it might behave in the future.
Forest Insect Management
In this course, you will have the opportunity to look at many different groups and types of forest health agents. The agent, the host, and the surrounding environment all interact, and each affects the other in various ways (Fig. 12). As you gain experience with the major forest insects of BC, keep this triangle in mind. Good forest management examines ways to manipulate the host, the environment, and/or the agent, rather than simply attempting to control the agent. Strategies (overall approaches), and tactics (specific methods within those approaches), must be considered within the context of objectives and values. It is also important to assess the efficacy of those strategies and tactics, and modify them as necessary.
