Lecture Materials
Bark Beetles Overview
Bark Beetle Ecology and Management
Introduction
Bark beetles are among the most economically important insects in forests. https://www.nbclearn.com/portal/site/learn/freeresources/changing-planet/cuecard/54762. They are insects, so they are in the phylum Arthropoda, class Hexapoda (or Insecta), and the order Coleoptera, the beetles. Some years ago, bark beetles were in a separate family called the Scolytidae, but they are now in the subfamily Scolytinae of the family Curculionidae, or weevils. Most weevils have a long snout, which bark beetles lack but, genetically, bark beetles are simply specialized weevils. The ambrosia beetles are also in the Scolytinae; these are specialist wood-boring beetles that feed on symbiotic fungi.
Table 1: Classification of bark beetles
|
Taxon |
Name |
|---|---|
|
Phylum |
Arthropoda |
|
Class |
Hexapoda (Insecta) |
|
Order |
Coleoptera |
|
Family |
Curculionidae |
|
Subfamily |
Scolytinae |
General biology
Bark beetles feed within the subcortical tissues of trees at some point during their life cycles. They may be monophagous or oligophagous. Some specialize in feeding on softwoods, while others specialize on hardwoods. There are about 6000 species worldwide, mostly in tropical and subtropical areas. There are about 480 species in the United States, and about 180 species in Canada.
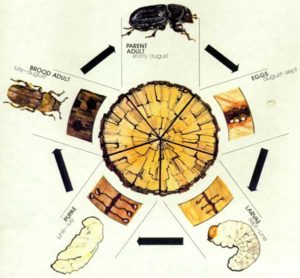
Bark beetles, like all beetles, are holometabolous, developing through egg, larva, pupa, and adult stages. Almost the entire life cycle is spent in/on a tree. Adults disperse to new host trees by flying, and may feed on the tree where they emerge, or on new trees. Following flight, they colonize new hosts and reproduce. A full life cycle, from emerging adult to the next generation of emerging adults, can take months to years. The emergence and dispersal part of that life cycle may only last for days to weeks.
Reproduction
Once a bark beetle finds an acceptable new host tree, it will initiate a gallery under the bark, excavating the subcortical tissues in a pattern that is characteristic of the beetle species on a particular host. The gallery is also inoculated with fungal spores.
Some bark beetles, e.g. Ips species, are polygamous: the male initiates the gallery, and attracts several females. After mating, the females begin excavating egg galleries, moving out from the central space (called the nuptial chamber) in different directions. This results in a characteristic pattern of egg galleries radiating out from the central nuptial chamber. https://www.flickr.com/photos/151887236@N05/41559658490. Other bark beetles, e.g. Dendroctonus species, are monogamous. In these beetles, the female initiates the gallery, and is later joined by a single male beetle. The pattern of galleries in these beetles consists of a long, central egg gallery, with larval galleries extending perpendicularly. Used in combination with identification of the host tree, a bark beetle gallery pattern can be used as a sign.
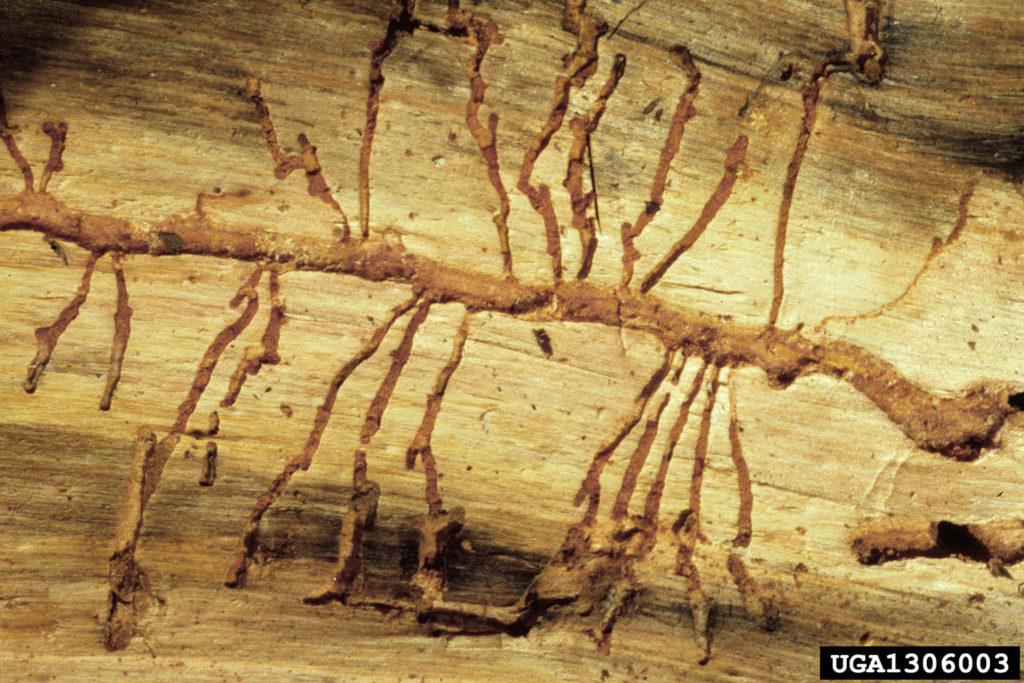
Female bark beetles oviposit (lay) less than 300 eggs each, with the number depending on the beetle species and individual condition. These may be deposited singly in egg niches along the length of the main gallery, or in groups. Males assist females with the excavation of the galleries, and have several adaptations for clearing frass and boring dust out of the galleries. For example, the frons, or front of the head, is often flattened, as are the tibiae of the legs. Bark beetles in the genus Ips have an elytral declivity, or a flattened area on the tips of the elytra, which can be used to push frass and dust. Finally, viewed closely, bark beetles tend to have hairy bodies, which helps them to move sawdust around the galleries.



Once the eggs hatch inside the galleries, the larvae feed on the phloem of the tree (in most bark beetles). They develop through 2-4 larval instars, and pupate in niches at the ends of the larval galleries. When they emerge as adults, they are often pale and relatively soft at first, and their reproductive organs are not fully developed. At this stage, they are referred to as teneral or callow adults. In order to become reproductive, most bark beetles must feed on a host tree, a process called maturation feeding. In some species, maturation feeding occurs under the bark of the tree as a teneral adult. Other species will fly to a new host and feed on live bark or foliage.
Bark beetles, like all insects, are profoundly affected by temperature. Prevailing temperatures govern the number of generations that can develop in a single year, or voltinism. Semivoltine bark beetles take more than one year to complete their development. Univoltine bark beetles have one generation per year, while multivoltine beetles can complete multiple generations in a single year. It is important for the beetles that synchrony of emergence is maintained. They may not live very long as adults, so need to be able to find mates within a short window of opportunity. The insects may overwinter in the host tree, or in litter or soil on the forest floor. Overwintering beetles can be extremely cold-tolerant.
Emergence/dispersal
Emergence from the original tree may occur before or after overwintering, depending on the beetle and the climatic regime. Weather factors, such as temperature, humidity, and wind, influence bark beetle flight. There is usually a range of optimal conditions under which adults will fly, and this range is usually species-specific. For many species, temperatures need to be >16℃, and even low levels of precipitation or wind can be problematic for an insect that is ~6-7 mm in size. The timing of flight is critical, if an individual is to encounter potential mates and susceptible hosts, while avoiding predation and competition. Dispersal can also be risky, especially in patchy, heterogeneous habitats.
Short range dispersal involves active flight, and uses energy. It is generally used for finding new, susceptible host trees, and potential mates, and the majority of any population likely only disperses for short distances. A relatively small percentage of a bark beetle population may also disperse long distances. They are initially photopositive, moving towards light, then later become host-positive, orienting to the scent of host trees. Long-distance dispersal is a poorly understood part of bark beetle biology, but it can be important. It was instrumental in the dispersal of mountain pine beetle from British Columbia, across the Rocky Mountains, and into Alberta.
Following its arrival among some potential hosts, a beetle must select a tree in which to initiate its gallery and reproduce. There is plenty of debate about the mechanisms that guide this choice. Landing on trees at random could play a role, but so could a variety of sensory cues. For example, like many insects, bark beetles have sophisticated sense of smell, and the primary attraction to a host is likely due to host odours. It is also probable that different cues play roles at different stages in the host selection process, and that the physiological state of the individual beetle influences the relative importance of these cues.

Colonization
Once it has landed on a tree, the beetle must accept it as a host and attempt to bore into the bark, or leave and try to find another host tree. The first beetles to arrive and attempt to colonize a host tree are known as “pioneer” beetles. As they bore into the bark and reach the subcortical tissues, they release secondary attractants to draw in other individual bark beetles of the same species. These secondary attractants are produced by the insects, and are often metabolized from compounds in the tree.
Trees are not defenseless. They produce copious amou-

nts of resin, and multiple chemical compounds, in an effort to repel the beetles. Secondary bark beetles are those that tend to attack trees that have impaired defenses, usually because they are weakened by abiotic or biotic stresses, or are diseased or dying. Primary bark beetles are those that attack relatively healthy trees. Whether a beetle is behaving as a primary or secondary bark beetle has less to do with the species of beetle, however, and more to do with the size of its population. Mountain pine beetle, for example, is usually a secondary bark beetle, attacking dying, diseased, or stressed trees. It is only when populations reach high levels that the pioneer beetles are able to attract enough other beetles that they can overwhelm a healthy tree’s attempts to defend itself.
Interactions with other organisms
From the point of view of a bark beetle attempting to colonize a tree, any tree that is not well-defended will be easy to colonize but, chances are, many bark beetles will be attempting to use it as a host. On the other hand, a vigorous host will have fewer competitors under the bark, but will be much more difficult to colonize. There is a trade-off, then, between interspecific competition and host defenses exerting selection pressure on bark beetles (See Fig. 1 in Lindgren and Raffa 2013 http://www.entomology.wisc.edu/raffa/Research/AllPubs/Bark%20Beetles%20and%20Associates/Social%20Behavior/Lindgren%20and%20Raffa%20TCE%20Evolution%20of%20tree%20killing.pdf). A few bark beetles are able to survive and reproduce in live hosts, but most kill the host.
Aggregation pheromones, which attract other beetles of the same species, play an important role in overcoming the tree’s defenses. If enough beetles attack an individual tree, it will eventually exhaust its resources and be unable to defend itself any longer. These mass attacks allow primary bark beetles to kill even healthy, vigorous hosts. There is a trade-off, however: more beetles attacking means more competition for their offspring. In addition to aggregation pheromones, each beetle also produces a small amount of an anti-aggregation pheromone, which acts to repel beetles of the same species. When the density of beetles rises to a certain point, the amount of anti-aggregation pheromone reaches a threshold level that r

epels any new beetles from attacking the tree. They frequently come close, but then start attacking neighbouring trees instead.
Bark beetles are not alone when they invade a host tree. They may attack in large numbers, of course, but they also come with a range of symbionts including fungi, nematodes, bacteria, and various animals that use phoresy (“hitch-hiking” on the bark beetles) to disperse to new trees. Fungi are extremely important in bark beetle biology. The spores of symbiotic species are carried by the bark beetles in specialized structures called mycangia. Their growth inside the new host tree can help to block translocation of water and nutrients, weakening the tree and helping the beetle to invade successfully, or even killing the tree outright.
A wide range of organisms preys on or parasitize bark beetles. Vertebrates, such as woodpeckers, are able to extract the larvae from under the bark. Insect predators, such as checkered beetles, live under the bark, or walk around the surface of the bark hunting for beetles. A variety of parasitic wasps are able to locate bark beetle larvae from the outside of the tree, then lay eggs inside those larvae using long ovipositors. These natural enemies can cause substantial bark beetle mortality in some situations, but most estimates put mortality at 50% or less.
Epidemiology/population dynamics
Abiotic factors are generally the largest cause of mortality for bark beetles. In particular, temperature has both direct and indirect effects on survival and reproduction. Bark beetles are often well-adapted to cold temperatures during the overwintering phases of their life cycles. They enter a physiological state known as diapause, in which development is suspended. They may also develop tolerance to cold, building up cryoprotectants, compounds that lower the freezing point of their body fluids. Nevertheless, beetles can be killed by sudden or prolonged cold temperatures. Indirectly, temperature anomalies can disrupt the synchrony of emergence among the population, or affect the time of emergence so that more competition or tree defenses are encountered.
Bark beetles such as the Dendroctonus species undergo some relatively predictable population phases over time. These phases are characterized by factors such as the condition of the host trees attacked, attack densities, the role of secondary species (competition), the impact of mortality, and the mechanisms of population regulation. There are four distinct population phases: Endemic = normal state; incipient; epidemic; and post-epidemic (Safranyik and Carroll 2006; https://cfs.nrcan.gc.ca/publications/download-pdf/26039).
The endemic population phase is the usual state of most bark beetle populations. The population density is low, meaning that mass attack is not feasible. Emerging beetles attack small diameter trees with weakened defenses. There are many competitors in these relatively poor-quality hosts, and reproductive success is low.
Sometimes environmental factors provide an opportunity for populations to increase. There may be an increase in secondary beetle species, or anomalous weather patterns, either of which can weaken a greater number of trees. There is either reduced stand resistance to attack, or reduced environmental resistance, or both. The trees attacked by the bark beetle we are interested in still tend to be small and suppressed, but there are more of them, and the population of beetles starts to increase. Tree mortality during this endemic-incipient transition is scattered. The beetle population is very unstable, largely regulated by competition, and can often move back to endemic levels.
If the conditions favourable to this bark beetle persist, however, populations continue to increase, allowing the adults to access larger-diameter, better defended trees, while they also experience less competition from secondary bark beetle species. On average, beetles will have higher reproductive success in trees with greater phloem thickness. Populations can now move from the incipient phase to the epidemic phase.
Epidemic bark beetle populations attack healthy trees. The attack rate is high, and tree mortality is widespread. Reproductive success among the beetles is high, making their population resilient to very high rates of mortality. Population regulation is no longer dependent on competitors, but is regulated bottom-up by climatic factors and the availability of susceptible hosts.
The epidemic population phase is ended either by weather conditions that cause dramatically high bark beetle mortality, or by a lack of susceptible host trees. In the post-epidemic phase, most host trees are dead, and attack densities are still high but decreasing. There may be increasing competition from secondary bark beetle species, whose populations have built up by using the dead hosts left by the primary bark beetle. The impact of various mortality factors is population-dependent but, in general, the population is decreasing and unstable.
Major Species in BC
Dendroctonus ponderosae, mountain pine beetle
Dendroctonus rufipennis, spruce beetle
Dendroctonus pseudotsugae, Douglas-fir beetle
Dryocoetes confusus, western balsam bark beetle
presence of agent itself
modified, hardened forewing of the beetle
number of broods/generations of an organism in a year