Chapter 4: Components of Adaptive Immune System
Learning Objectives
By the end of this chapter you will be able to:
-
Identify the key components of the lymphatic system and describe the major functions of primary and secondary lymphoid organs/tissues
- Define and describe the clinical significance of immunogens, antigens, epitopes and haptens.
- Explain the differences between auto-antigens, allo-antigens and heterophilic/heterogenic antigens, with examples.
- Define Human Leukocyte Antigens (HLAs)/ Major Histocompatibility Complex (MHC).
- Describe the functions and tissue distribution and structure of Class I and II MHC molecules.
- Define haplotypes, isotypes, allotypes and polymorphism, in the context of HLA.
- Explain the inheritance of MHC genes and discuss their clinical significance.
Case Study
Daniel introduces himself as a 50 year old male who has a long history of alcohol abuse, resulting in cirrhosis (scarring) of the liver. Daniel was placed on a waiting list for a liver transplant and received a transplant after seven months. The surgery was successful, and initially, his new liver functioned well. After the transplant, Daniel was able to manage his alcohol addiction but still developed dark urine, jaundice and abdominal discomfort. A blood test found circulating liver enzymes indicating liver damage. Further investigation found that Daniel was experiencing transplant liver rejection. Following a long-term treatment with immunosuppressive medications, rejection of the liver subsided and Daniel benefitted from a functional liver.
- Where did the transplant come from and what kind of antigen might be found on the transplanted tissue?
- Which antigen is the most relevant in transplant immunology and why?
- Why did it take so long to match Daniel with a donor liver?
- What is the purpose of immunosuppressive medication in transplant rejection?
Answers to these questions are at the end of the chapter.
4.1 Overview of Adaptive Immune Response
For a vast majority of insults to the body, the innate immune response is sufficient to prevent infectious disease. As a consequence, we can be routinely exposed to potential pathogens without ever experiencing the effects of illness and infectious disease. However if an infection persists, a more strategic response to infection is engaged. This is called the adaptive response (aka the acquired response)
The adaptive response to infection extends from the innate response. Monocytes arrive at infected tissues and differentiate into macrophages or dendritic cells. These newly arriving cells support the existing neutrophils in phagocytizing pathogens. Monocyte-derived cells have an additional characteristic that neutrophils do not possess. These cells display fragments of the consumed pathogen, called antigens, on their surface. Antigens are presented to T-lymphocytes, which subsequently coordinate a distinct immune response that specifically targets this pathogen.
The adaptive immune response takes two basic forms: (1) the humoral adaptive response and (2) the cell mediated adaptive response. The humoral adaptive response involves producing antibodies that mark pathogens and hyperstimulate the immune system to eliminate them. The cell mediated adaptive response involves activation of macrophages and ‘cytotoxic’ T-lymphocytes that collectively specialize in eliminating pathogens that reside inside human cells, like viruses or intracellular bacteria.
The final phase of the adaptive response is the formation of memory lymphocytes. Memory lymphocytes are long-lived cell populations that enhances the rapidity and intensity of the immune response upon subsequent exposure to a pathogen in the future. Often, this future secondary immune response is extremely fast and intense that a person experiences little to no symptoms of the disease. This phenomenon is referred to as acquiring immunity to infectious disease.
4.2 Tissues of the Adaptive Immune Response
Leukocytes are dispersed throughout the body by the bloodstream and enter into the tissues by deforming through gaps between the endothelial cells that line the capillary vessels. Leukocytes then migrate through the tissue fluid, called the interstitial fluid, and respond to any pathogen within the tissue space.
Leukocytes are then drained, along with tissue fluid and debris, into vessels called lymphatic vessels (also called lymph vessels). Interstitial fluid is filtered through lymph nodes, distributed around the body, before being return to the bloodstream. Lymph nodes are one of the lymphoid organs and are packed with lymphocytes, including T-cells and B-cells.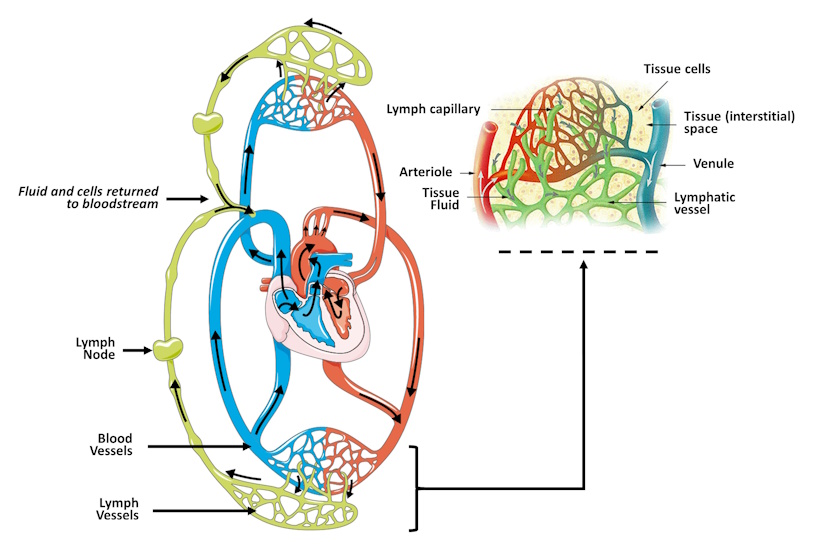
Figure 4-1 Lymphatic drainage. Fluid and leukocytes from the bloodstream flow into the interstitial space within the tissues. Fluid that does not passively return to the bloodstream is drained into local lymph capillaries. These fluids, along with tissue debris and potential pathogens, are filtered through lymphocyte-rich lymph nodes before returning to the bloodstream. [Link to Image Description]
Image Source: Main image by Alicia Fagerving CC BY 3.0, via Wikimedia Commons; Inset by Lennert B CC BY 3.0, via Wikimedia Commons
Lymphatic organs and tissues are classified into two categories: (1) primary lymphoid organs where immune cells are produced and (2) secondary lymphoid organs where immune cells are stored. Primary lymphoid organs include bone marrow and thymus in adults, which replace the yolk sac and liver as the sites where immune cells are produced and matured after birth. Secondary lymphoid organs include lymph nodes, spleen, tonsils and mucosa-associated lymphoid tissues (MALT).
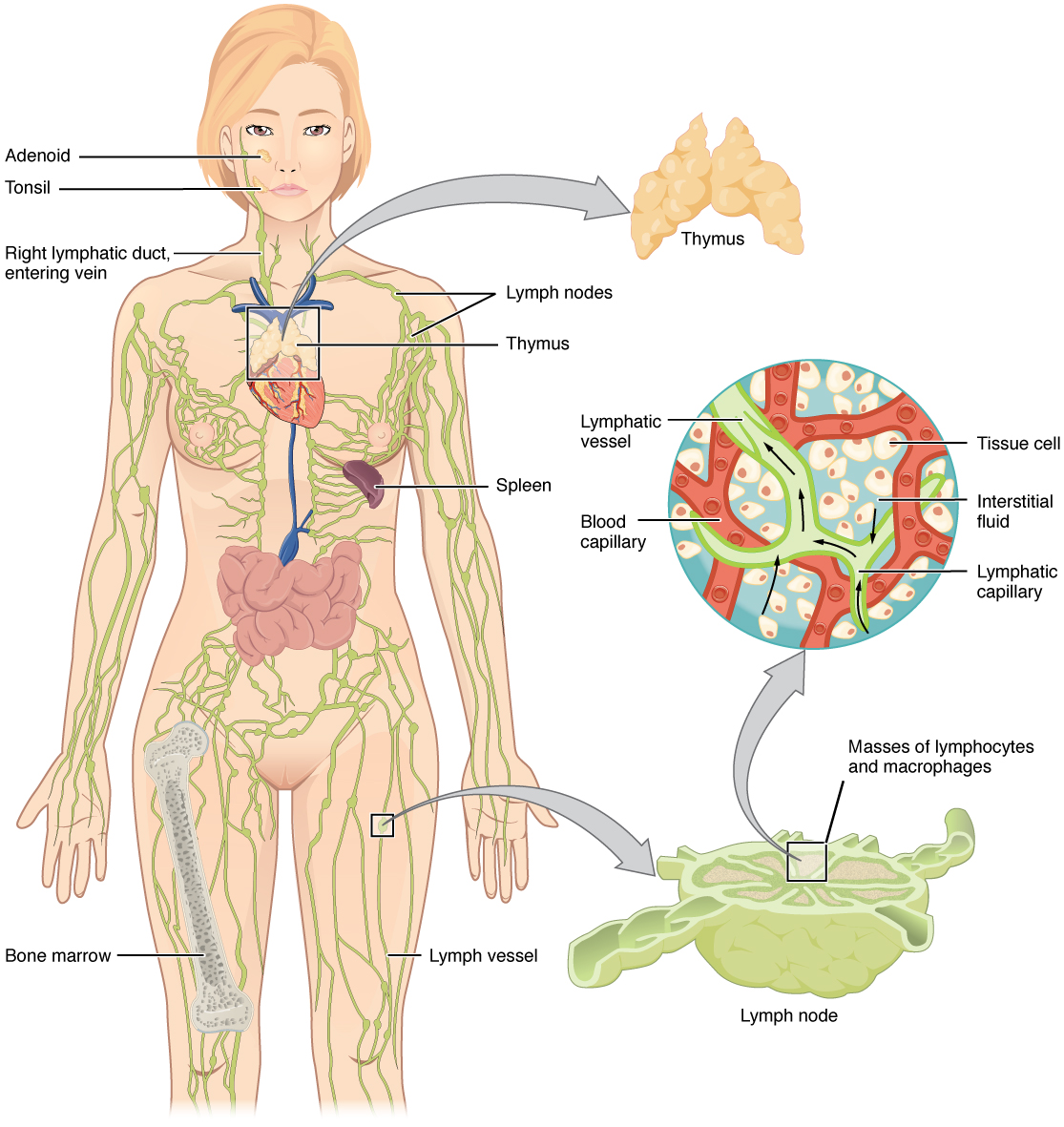
Figure 4-2 Lymphoid organs. Lymphoid organs are distributed throughout the body and play distinct roles in preventing infection within sensitive tissues. [Link to Image Description]
Image Source: By OpenStax CC BY 3.0, via Wikimedia Commons
The functions of lymphoid organs
Bone marrow: Primary lymphoid organ responsible for hematopoiesis, the process of forming red blood cells, white blood cells and platelets within the red bone marrow in the core of the bone.
Thymus: Primary lymphoid organ where immature thymocytes mature into functional T-cells. Also the site of T lymphocyte selection to ensures that mature T-cells do not respond to human tissues.
Lymph node: Filters interstitial fluid before it returns to the circulatory system. Lymph nodes are also the site where antigen is presented to B-cells and T-cells to produce an adaptive immune response when pathogens are encountered in the tissues.
Spleen: Filters blood during routine circulation. Rich in macrophages and dendritic cells that remove pathogens and dying red blood cells from the bloodstream. Infections within the bloodstream pose a serious risk, making the role of the spleen extremely important.
Tonsils: Lymphoid nodules that survey microbes in the upper digestive and respiratory tracts. Swollen tonsils can be an important sign of infection.
Mucosa-associated lymphoid tissue (MALT): Various tissues throughout the body that protect regions that are particularly vulnerable to infection. These tissues vary in name by location (e.g. GALT=Gastrointestinal; BALT=Bronchus airway). The Peyer’s patch is an important MALT of the small intestine. MALT can also be found within the appendix. Intestinal blockage may lead to inflammation of the appendix, resulting in painful appendicitis.
4.3 Antigen
Recall that phagocytes detected pathogens based on general patterns (PAMPs) commonly found on the surface of multiple microbes (e.g. a cell wall is common to most bacteria). Conversely, the adaptive immune system responds to antigens, which are often specific to a particular pathogen. For example, a person may be exposed to influenza antigen to stimulate immunity through vaccination. However, these antigens are no found in other viruses as such the influenza vaccine will not protect against other viruses.
Antigens were traditionally named because they were capable of inducing antibody generation. However, it is now known that the adaptive immune response includes more than just antibody production. According to the current understanding antigen represents specific molecules that can be bound by specific antibodies or lymphocyte receptors (T-cells and B-cells). However, binding antigen alone is not always sufficient to trigger a productive adaptive immune response. Antigens that generate an adaptive immune response are called immunogens.
Antigenicity
Another important concept related to antigens is the typical intensity of antigen binding (antigenicity), which often relates to the intensity of the induced response (immunogenicity). Antigenicity depends largely on the three-dimensional of the molecule. Of particular importance is the precise part of the molecule where the antibody or lymphocyte receptor attaches designated the epitope. Epitopes are very small regions of the protein (as small as 4-6 amino acids) that adopt a unique three dimensional conformation. Owing to their role in antigen binding, epitopes are also called antigenic determinants.
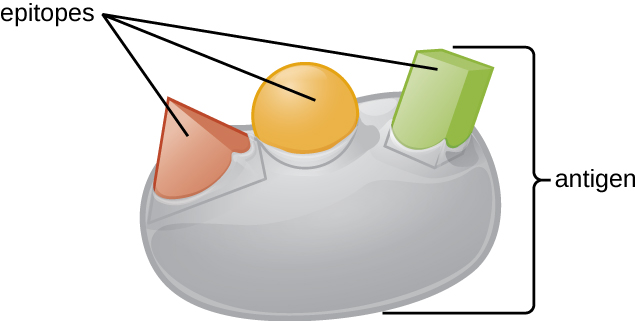
Figure 4-3 Antigen. An antigen is a specific molecule with one or more defined three-dimensional shapes, called ‘epitopes’. This image illustrates one antigen with three different epitopes. [Link to Image Description]
Image Source: By OpenStax CC BY 3.0, via Wikimedia Commons
Various characteristics of the antigen are important in determining its intrinsic antigenicity.
Size of the antigen is important because the substance must be large enough to physically interact with immune cells. Typical protein units (peptides) range 10-30 kDa in size. While antigenic molecules less than 10 kDa in size are only weakly immunogenic. For example, the molecules in perfumes and scented detergents are so small that they rarely trigger new immune responses. However, once a person develops an adaptive immune response toward these perfumes this response may be trigger whenever the molecules are encountered, in the form of an allergic response.
- If small molecules are poorly immunogenic, how do people develop adaptive responses (including allergies) toward them? Small antigens that are antigenic but not immunogenic are called haptens. By attaching haptens to larger molecules, called carrier molecules, an immune response can be triggered against the hapten-carrier complex, which will potentially provide long term immunity. This strategy is used in development of some vaccines. For example, Haemophilus influenzae bacteria have a capsule coat made of sugars too small to serve as vaccine immunogens. Physically attaching (conjugating) these sugar molecules (hapten) to larger inactivated tetanus toxin molecules (carrier) produces a new conjugate molecule that is sufficiently immunogenic for use in vaccines.
Proteins are typically more immunogenic than polysaccharides (sugars) or lipids (oils and fats). However, the other characteristics of the molecule may enhance the immunogenicity of polysaccharides and even lipids.
Complex molecules with several distinct epitopes are more likely to be immunogenic than microbes with few epitopes or molecules that simply consist of a long chain where the same epitope structure is repeated. For example DNA and RNA are large molecules bound by proteins but their regular repeating structure ensures that they are less frequently able to trigger an adaptive response.
Accessibility of epitopes is often difficult to predict because molecules may form complex three-dimensional structures. However, immunogenic antigens typically have the relevant epitopes on the outer surface, where they are more readily detected by immune cells.
Foreignness of the molecule within the human body is a consideration in its immunogenicity. The adaptive immune system has a mechanism for tolerance for human cell antigens as well as antigens that frequently occur within the microbiome.
- As is the case with haptens, immune tolerance only makes human-like antigens less immunogenic. These molecules may still be antigenic once the adaptive response is initiated. This is exemplified in the context of molecular mimicry, where a microbe may have antigen structures that mimic human antigen. The outcome of molecular mimicry is that the immune response toward a pathogen may cross-react to bind unrelated proteins on human tissues. Streptococci bacteria have a surface protein called the M-protein, which has similar epitope structure to human mysoin/tropomysin found in heart muscle. While these epitopes are poorly immunogenic, during a severe infection the adaptive response may be triggered to react toward them. Once the adaptive response is triggered, antibodies will continue to be produced against the M-protein and cross-react to also bind proteins in heart muscle. The outcome is that the streptococcal infection will be effectively cleared but the antibodies will produce chronic illness, presenting as carditis and rheumatic fever.
Classification of Antigens
Autoantigens represent the body’s self-recognition system. These antigens emerge from an individual’s own tissues and cells, reflecting the inherent distinction between the body’s components and potential threats. As such, cells of the adaptive immune system undergo a strict selection process to ensure that they do not react significantly to these ‘self’ antigens. In many cases, self-reactive cells are killed and deleted during maturation. Alternatively, the lymphocyte receptor that dictates the specificity antigen recognition may be modified to no longer detect self-antigen, or the cells may be regulated to reduce the amount of this receptor protein on the cell surface.
However, in certain circumstances, the immune system can erroneously trigger adaptive responses against autoantigens. The example of streptococcal M-protein and human myosin cross-reactivity, described above, is an example of such an erroneous event. The outcome of adaptive response to self-antigen is autoimmune disease, which can manifest in different ways depending on the localization of the autoantigen in the body. For example, autoimmune reactions against the human pancreas can lead to destruction of the tissues that produce and secrete insulin, resulting in type 1 diabetes.
Alloantigens reflect the diversity of each person’s unique genetic identity. Alloantigens are differences in antigen structure between individuals within the same species. While variation may occur in virtually every protein that is expressed by our cells, the most significant variation is found in the protein structures that make up the major histocompatibility complex (MHC). The MHC plays a pivotal role in immune detection of pathogens (described later). To prevent microbes from adapting to overcome detection, the MHC has evolved extensive variation such that identical matches between any two people by chance alone is highly unlikely. The significance of alloantigens lies in their involvement in transplantation and transfusion reactions. When the immune system encounters foreign MHC molecules in transplanted organs or transfused blood, they might be perceived as non-self evoking a rigorous immune response. The result of this immune response is that the transplanted tissue is rejected and destroyed by the recipients immune system.
Heterophilic antigens bridge the gap between species. Heterophilic antigens occupy a unique niche in immunology by fostering connections between different species. The heterophilic antigen has at least one epitope that is found in one species but can induce an immune response when encountered by the immune system of another species through cross-reactivity. This effect is directly exemplified by the streptococcal M-protein and human myosin cross-reactivity, described above. In medical laboratory testing, heterophilic antigens can lead to false diagnoses that might profoundly impact clinical decision and the welfare of people who rely on them. For example, testing for the syphilis, a sexually transmitted infection (STI), may rely on detecting antibodies against cardiolipin, a component of the bacterial membrane. However, cardiolipin is also found in the mitochondria of human cells and may be externalized in some severe conditions, such as lupus. As many as 50% of lupus patient will make antibodies against the heterophilic cardiolipin antigen and will test positive for syphilis even though they do not have the STI.
4.4 Lymphocytes and Lymphocyte Receptors
Lymphocytes are a subset of the leukocytes (white blood cells) that include B-cells, T-cells and natural killer (NK) cells. NK cells are similar to cytotoxic T-cells, but kill target cells following detection of PAMPs and DAMPs, making NK cells components of the innate immune response. This chapter will focus on the T-cells and B-cells as lymphocytes responsible for the adaptive immune response. T-cells and B-cells have cell surface proteins called lymphocyte receptors that are structurally different but function similarly in determining the specific antigen that these cells will respond toward.
Each B-cell and T-cell develops from the common hematopoietic stem cell and mature into functional lymphocytes. During maturation, they develop lymphocyte receptors that are unique in shape from one cell to another. The mechanism by which this occurs will be described in Chapter 5. The outcome of this maturation process is that populations of T-cells and B-cells emerge, called clones, each containing a unique receptor that will bind to only a single epitope. B-cells have the ability to secrete their lymphocyte receptor (B-cell receptor, or BCR) as an immunoglobulin that acts as an antibody in the immune response. Immunoglobulins and antibodies will be discussed in greater detail in Chapter 5. In Figure 4-2, we observe that three clones of B-cell have independently produced antibodies. Since the BCR of the originating clone was different, the shape of the secreted antibodies are also sufficiently different that they bind only to their matching epitope.
Since antigens almost invariably have more than a single epitope, the antibodies produced against them are a mixture of antibodies from multiple different B-cell clones. As a consequence, the antibodies produced in a natural immune response are called polyclonal antibodies. In the laboratory, it is possible to isolate single B-cells and to form a clonal population of cells , which produce the same antibody that all bind to only a single epitope. These laboratory-derived antibodies are called monoclonal antibodies.
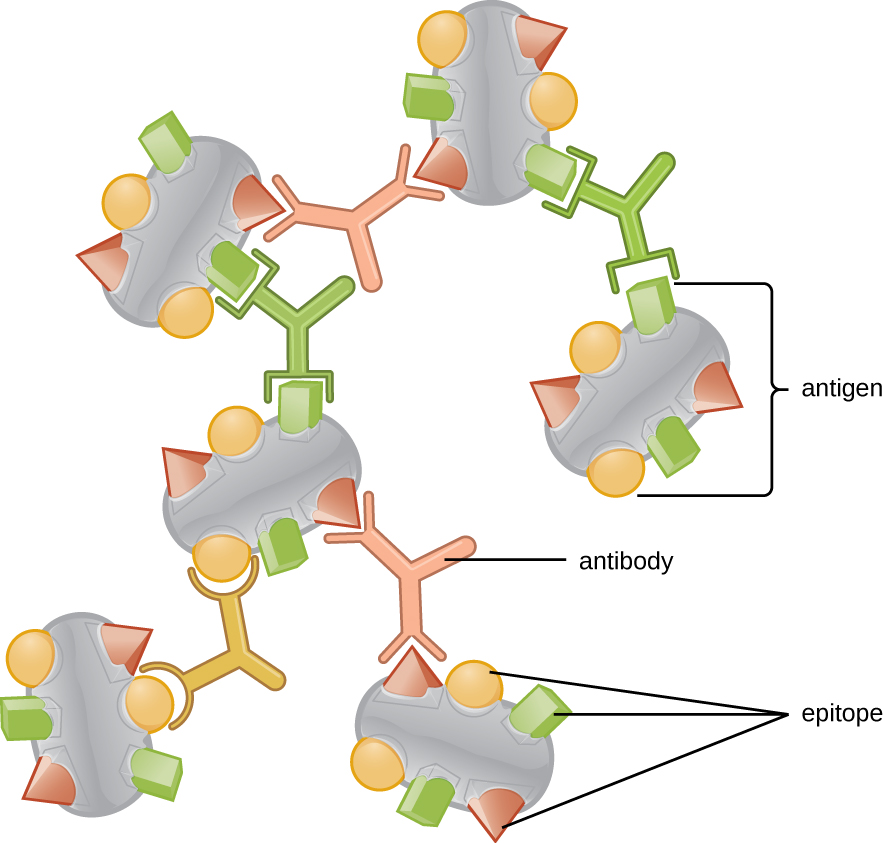
Figure 4-4 Polyclonal Antibodies. Antibodies are B-cell receptors that are secreted into blood and other tissues. Each B-cell clone has a receptor for a single epitope. Since antigens have multiple epitopes, they typically trigger the production of antibodies from multiple B-cell clones, making the antibody mixture polyclonal. [Link to Image Description]
Image Source: By OpenStax CC BY 3.0, via Wikimedia Commons
4.5 Antigen Presenting Cells
In the previous section, we introduced clones of B-cells and T-cells each binding and responding to different epitopes. Once these cells respond to the bound antigen, they set in action a cascade of events that may produce a longterm immune response toward the antigen, commonly known as ‘immunity’. However, the adaptive immune response is not triggered by antigen freely floating in the interstitial space between cells. Instead, there are a special subset of phagocytes called antigen presenting cells (APCs) that display potential antigen molecules on their surface and assist in the initiation of the adaptive immune cascade. These antigen presenting cells include macrophages, dendritic cells and B-cells.
In a standard immune response, macrophages and dendritic cells phagocytose, degrade and process pathogen cells and debris, and then present the molecular fragments on their surface to elicit a response from an important T-cell subpopulation called helper T-cells. The helper T-cells then activate other T-cells and/or B-cells to produce a tailored and appropriate response to the infectious agent. This antigen presentation process is performed by a large molecular complex called the major histocompatibility complex, type 2 (MHC-II).
Like macrophages and dendritic cells, B-cells are also APCs. However. unlike macrophages and dendritic cells, B-cells only phagocytize cells or debris that have a matching antigen to the BCR. B-cells then present internalized antigen to already activated helper T-cells and the B-cell becomes activated in response to this interaction.
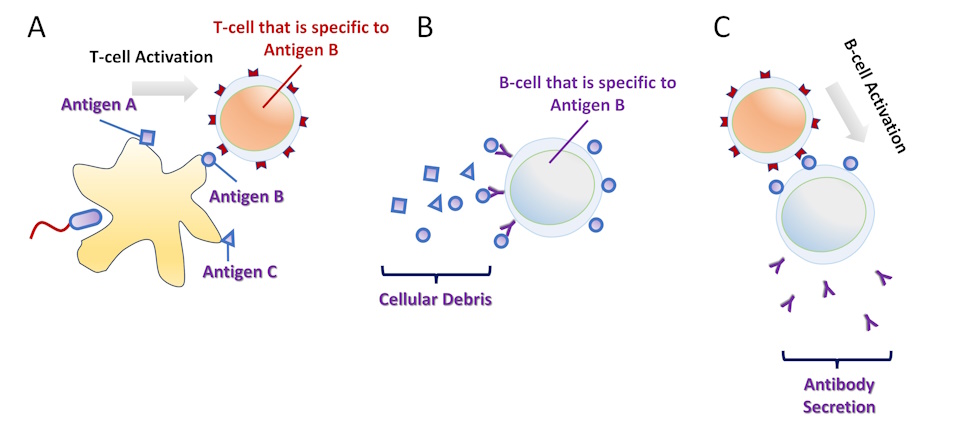
Figure 4-5 Antigen Presentation. (A) A macrophage or a dendritic cell phagocytize a pathogen and presents its antigens on the cell surface. A helper T-cell that is specific to one antigen binds the APC and becomes activated. (B) Independently, cellular debris flows in the vicinity of a B-cell. The B-cell receptor is specific to one antigen and this antigen is phagocytized and presented by the B-cell. (C) When there is contact between an activated helper T-cell and a B-cell recognizing the same antigen, the B-cell becomes activated and release their antigen-specific B-cell receptor as antibodies. [Link to Image Description]
A secondary form of antigen presentation involves the major histocompatibility complex, type 1 (MHC-I). This process is performed by all nucleated cells, not just by phagocytes. Proteins that are produced within the cell are routinely shuttled to the cell surface and presented by MHC-I. Abnormal proteins, such as those produced as a result of viral infection or within a tumor cell may be detected by cytotoxic T-cells. While helper T-cells function by triggering a broader immune response to an antigen, cytotoxic T-cells simply kill the cell that presents the foreign antigen. The cytotoxic response is an effective way to prevent the spread of infection from one cell to another. However. in the context of a large-scale or chronic infection, the cytotoxic response can kill cells that perform an important function and thereby severely damage the human tissue.
A classic clinical example where cytotoxic T-cells severely damage human tissue is seen in organ transplant rejection. When an individual receives an organ transplant, such as a kidney or heart, from a donor, the immune system of the recipient recognizes the transplanted organ as foreign due to differences in the major histocompatibility complex (MHC) molecules themselves. The cytotoxic T-cells will destroy the grafted tissue because of these foreign molecular markers and reject the transplanted tissue.
The adaptive immune response begins with phagocytosis by macrophages and dendritic cells. APCs will process the phagocytized pathogen and present it on the cell surface using a molecular complex called the major histocompatability complex (MHC).
MHC-I – All nucleated cells present elements of the proteins they produce, using the MHC-I complex. If the cell is infected by a virus or develops into a tumor cell, the aberrant proteins produced from those events are presented on the cell surface to alert immune cells to this problem.
MHC-II – In addition to MHC-I presentation, some cells also present products of phagocytosis using the MHC-II complex. These professional antigen presenting cells include macrophages, dendritic cells, and B-cells. Other immune cells can present antigen to a lesser degree.
MHC and Tissue Transplantation
In transfusion medicine, the MHC is known by a different name: Human leukocyte antigen, or HLA. MHC-I and HLA-I are synonymous, as are MHC-II and HLA-II. All of the genes that make up the HLAs are clustered together on chromosome 6, and are often described as a “molecular barcode” that is virtually unique to every person.
- Human MHC-I consists of three genes, or isotypes, called HLA-A, HLA-B, HLA-C.
- Human MHC-II also consists of three isotypes, called HLA-DP, HLA-DQ and HLA-DR.
Recall that MHC-I antigens will be expressed on all nucleated cells, while MHC-II are mainly restricted to antigen presenting cells (monocytes, dendritic cells, macrophages, B-cells).
For each of these isotypes there are thousands of possible variations from one person to another, creating an incredible diversity of possibilities. As such, the possibility of perfectly matching all six isotypes between two individuals at random is virtually impossible.
Making a perfect match even less likely because the set of HLA-A, -B, -C, DP, -DQ and -DR on one chromosome constitutes only half of the entire repertoire, called a haplotype. Because of codominant genetics, a person inherits two distinct chromosomes. These chromosomes originate from each of the two parents. The result is that perfect matching would require a match of 12 different isotypes, where each isotype has thousands of possible variations.
In the context of a tissue or organ transplant from a donor to a recipient, it might seem virtually impossible to establish a match and avoid rejection. It should be noted, not all mismatches are equally deleterious.
- Different isotypes occur at different frequencies in human populations. Since HLA isotypes are inherited, these frequencies depend on a person’s ancestry.
- Not all HLAs equally immunogenic. The most immunogenic is HLA-DR, followed by HLA-B and -A. These 3 loci are therefore the most important for matching donor and recipient.
- Some mismatches are tolerated better than others because of a person’s intrinsic immune tolerance.
Consequently, determining the best donor-recipient match is a complex task but can have a significant impact on the success of a tissue or organ graft.
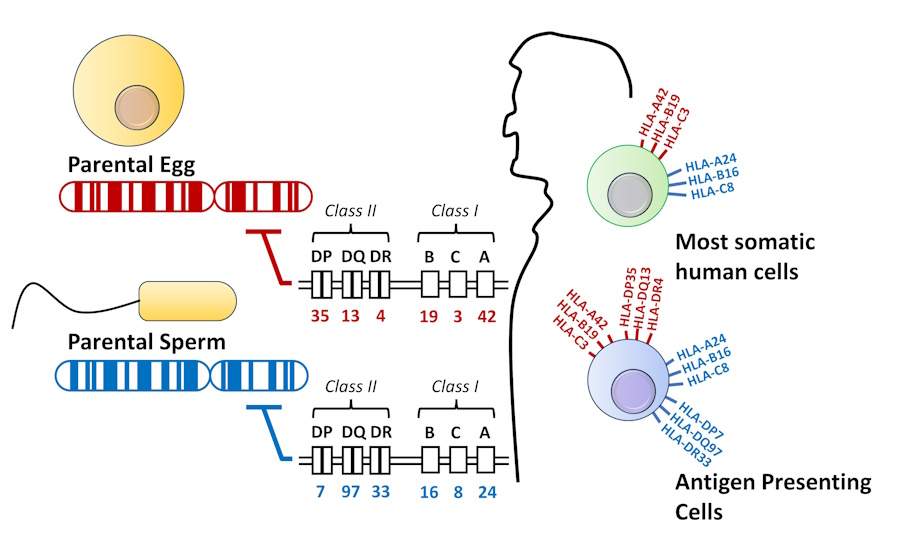
Figure 4-6 MHC Genetics. Class I and class II MHC/HLA are both encoded on chromosome 6. Each person receives two copies of this chromosome from the parental egg and sperm. Most somatic cells only express the class I MHC. Antigen presenting cells express both class I and class II MHC on their surface. [Link to Image Description]
4.6 Coordination of the Components of the Adaptive Immune Response
Each component of the adaptive immune response acts in a coordinated an orchestrated manner with one another.
The adaptive immune response emerges from a persistent innate immune response to infection, as shown in the series of events below.
- Persistent infection induces inflammation and recruitment of antigen presenting cells to the tissue, such as macrophages and dendritic cells.
- Antigen presenting cells phagocytose pathogen and debris, presenting the foreign material on the cell surface using MHC-II.
- Helper T-cells with T-cell receptors that match this particular foreign antigen will bind with the antigen presenting cells and signal a broader adaptive immune response.
- The adaptive immune response may include activation of cytotoxic T-cells, which scan all proteins presented by MHC-I on the surface of almost all cells in the body. If foreign protein is produced inside the cell (due to infection or cancer) that antigen will be detected and the aberrant cell will be killed.
- The adaptive response may include activation of B-cells, which release their B-cell receptors as antibodies to support in the defense against infection.
- One outcome of the adaptive immune response is that future infections will normally elicit a faster and stronger response, which we often regard as “immunity“
Summary
- The adaptive immune system recognizes antigens found on the surface of pathogens or foreign substances.
- Antigen recognition is mediated by specialized lymphocytes, primarily B cells and T cells.
- Antigen-presenting cells (APCs), such as dendritic cells, engulf and process pathogens, breaking them down into smaller antigen fragments.
- These antigen fragments are then displayed on the surface of APCs along with major histocompatibility complex (MHC) molecules.
- T cells are activated when they recognize antigens presented by APCs. There are two main types of T cells: helper T cells (CD4+) and cytotoxic T cells (CD8+). Helper T cells stimulate other immune cells, while cytotoxic T cells directly target and kill infected cells.
- B cells are activated when they encounter antigens matching their specific antibody receptors. This activation leads to the production of antibodies, proteins that can bind to and neutralize the pathogen.
- Human leukocyte antigen (HLA) play a pivotal role in organ transplantation.
- HLA molecules are responsible for presenting antigens to immune cells, particularly T cells. They serve as a bridge between the innate and adaptive immune responses.
- HLAs are highly polymorphic, meaning they exist in numerous different forms or alleles within the human population. This diversity allows the immune system to recognize a wide range of antigens.
- Matching HLAs between donors and recipients is critical for the success of organ and tissue transplantation. A close HLA match reduces the risk of graft rejection, as the immune system is less likely to recognize the transplanted organ as foreign.
Pathogen associated molecular patterns (PAMPs) are the general features that our innate immune system uses to identify foreign agents, like bacteria and viruses. PAMPs are different from "antigens", which are molecules that are specifically identified by cells and antibodies of the adaptive immune system.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}