Chapter 5: Development and Stimulation of Adaptive Immune Response
Learning Objectives
By the end of this chapter you will be able to:
-
Outline the series of events that occur during the adaptive immune response
- Identify the different T-cell populations and how they regulate the immune response
-
Describe the origin and development of T and B cells
- Discriminate between the humoral and cell mediated immune responses
-
Describe the cytotoxic mechanism of cytotoxic T-cells, and related lymphocytes
-
Explain the role of memory cells in protecting the body from reinfection
-
Distinguish between the T-dependent and T-independent humoral immune response
Case Study
After finishing college Bernice decided to experience international travel. As a health conscious traveller, Bernice received the necessary travel vaccines. Her vaccines included the hepatitis A vaccine, since this viral infection spreads from contaminated food and water in places throughout the world and can produce acute liver damage, leading to jaundice, nausea, vomiting and lethargy.
The hepatitis A vaccine involves injecting inactivated virus into the human body and using two doses. While travelling, Bernice was not exposed to hepatitis A. She returned home healthy and tremendously enriched by the experience. Five years later, Bernice consumed a raw oyster that was contaminated with hepatitis A. Bernice was infected by the virus but did not have any noticeable illness. Bernice’s immune system rapidly cleared the infection underscoring the significance of vaccination in bolstering the body’s immune repertoire and enhancing its ability to combat infectious diseases.
- Why does Bernice need travellers’ vaccines when she received multiple vaccines, including hepatitis A, as a child?
- When should Bernice get the vaccine, prior to travelling?
- Why did Bernice experience no symptoms when infected by hepatitis A virus from oysters? Could a vaccinated person develop symptoms of infection?
- What mechanisms allow the immune system to eliminate viruses, like hepatitis A?
Answers to these questions are at the end of the chapter.
5.1 Lymphocytes Diversity
As mentioned in the previous chapter, lymphocytes include T-cells, B-cells and natural killer cells. Lymphocytes can be further divided into cell subsets with distinct and important roles in the immune response. The most diverse lymphocyte is the T-cell. T-cells can be broadly divided into CD4 and CD8 cells, based on “clusters of differentiation” (CD) proteins on their surface.
CD4 binds MHC-II facilitating the previously described interaction between helper T-cells (CD4+) and antigen presenting cells (APCs, MHC-II+). When APCs present antigen to CD4 helper T-cells, those T-cells can stimulate a potent adaptive immune response. However, there are multiple types of CD4 cells that can produce a tailored immune response to different antigens as shown below. CD8 binds MHC-I and allows cytotoxic T-cells to perform surveillance on all nucleated human cells. If these CD8 cytotoxic T-cells detect foreign antigen on the surface of a cell, they presume the cell is infected and destroy it.
B-cells can also be subdivided based on the context of antibody secretion. The cells that secrete antibody are designated plasma cells. Alternatively, B-cells may form memory cells that contribute to longterm immunity.
T-cells have an antigen-specific receptor, called a “T-cell receptor” and participates in the cell-mediated adaptive immune response. Upon binding their antigen, T-cells may have different responses, depending on the type of T-cell:
- Type 1 Helper T-cell (CD4+ Th1) – Effector cells that secrete cytokines to stimulate an immune response. If human cells are infected by intracellular pathogens (e.g. viruses or bacteria), or have developed into tumor cells, the Th1 will stimulate cytotoxic T-cells to kill the infected cell. Th1 also stimulate innate immune cells, such as macrophages, neutrophils and NK cells.
- Type 2 Helper T-cell (CD4+ Th2) – Effector cells that secrete cytokines to stimulate an immune response. Coordinate the humoral response that involved B-cell differentiation and secretion of antibodies.
- Type 17 Helper T-cell (CD4+ Th17) – Effector cells that secrete proinflammatory cytokines to stimulate an immune response. Important in defense against extracellular bacteria and fungal of the skin and mucous membranes.
- Regulatory T-cell (CD4+ Treg) – Cells that regulate and often suppress the antigen-specific immune response. Secretes cytokines that promote tolerance to microbes that are normally resident to the human body and suppresses immune responses against the body’s own tissues (autoimmune responses).
- Cytotoxic T-cell ( CD8+ CTL or Tc) – These cells can bind MHC-I on the surface of nucleated cells and monitor antigens produced within the cell. If the cell has recognized abnormal antigen, indicative of infection or tumor aberrant protein expression, the cell is killed. The CTL releases granzymes to trigger apoptosis controlled cell death in the target cell as well as perforins to form pores that disrupt the cell surface and lyse (burst) the cell.
- Memory T-cell – Similar to memory B-cells, memory T-cells persist after the infection and result in a faster and more exaggerated secondary immune response, for future infections with the pathogen.
B-cells have an antigen-specific receptor, called a “B-cell receptor” and participates in the humoral adaptive immune response. Like monocytes, B-cells phagocytose pathogens with these antigens on their surface. Also like monocytes, B-cells present antigen on their surface. In contrast to monocyte antigen presentation, B-cells do not activate T-cells but instead become activated through this interaction. B-cells differentiate into plasma cells that secrete their B-cell receptor as antibodies. Some B-cells also differentiate into memory cells that persist after the infection and can stimulate a more effective secondary response upon future exposure to the pathogen.
Natural Killer (NK) cells are cells of the innate immune response and recognize PAMPs using cellular receptors like those found in neutrophils. Unlike neutrophils, NK cells are not phagocytes. They kill the target cell using the same mechanisms employed by cytotoxic T-cells.
Maturation of Lymphocytes
As with other white blood cells (leukocytes), B-cells, T-cells and NK-cells (lymphocytes) are produced through the process of hematopoiesis (see Chapter 2.3). B-cells and T-cells undergo further maturation, which involves the development of lymphocyte receptors, called B-cell receptors (BCR) and T-cell receptors (TCR). During maturation, these receptors undergo genetic V(D)J recombination to produce a unique receptor on each lymphocyte that is specific to a single target antigen (or few structurally related antigens).
V(D)J Recombination allows individual T-cells and B-cells to bind different antigens. Since lymphocytes are constantly being produced in large number, the combination of all of the antigens that can be bound by B-cells and T-cells is known as the immune repertoire. The BCR and TCR receptors are different in structure but are composed of multiple interconnected chains that are held together by disulfide chemical bonds.
- BCRs are made of four chains that include a pair of ‘heavy chains’ and a pair of ‘light chains’ that attach to form a characteristic ‘Y’ shape.
- TCRs are typically formed from two attached chains, called the alpha (α) and beta chains (β).
- In some T-cells, alternative gamma (γ) and delta (δ) chains make up the TCR.
| A. | B. |
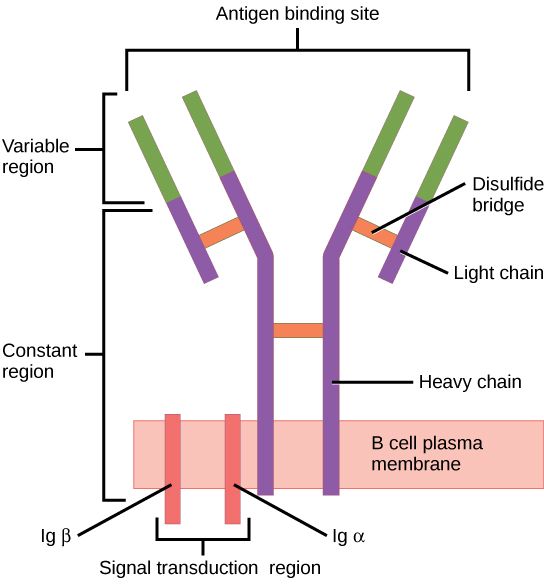 |
Figure 5-1 Lymphocyte Receptors. (A) B-cell receptor is a membrane bound immunoglobulin that consists of four polypeptide chains: two light chains and two heavy chains. This produces two sites within the ‘variable regions’ for antigen binding on each immunoglobulin molecule. [Link to Image Description] (B) T-cell receptor consists of two membrane-bound polypeptide chains that come together to form a single antigen-binding ‘variable region’. [Link to Image Description]
Image Source: (A) By OpenStax CC BY 3.0, via Wikimedia Commons; (B) By OpenStax CC BY 3.0, via Wikimedia Commons
V(D)J recombination is achieved using RAG proteins that remove DNA from the part of the lymphocyte genome that codes for the variable region of the lymphocyte receptor, which is the region that binds antigen. The diversity of these chains occur because there are many repeating copies of the relevant gene segments, which are named Variable, Diversity and Joining genes. The RAG proteins remove all copies except one of each gene segment.
- The BCR heavy chain as well as the TCR β-chain undergo VDJ recombination, where the peptide receives three randomly selected genes.
- The BCR light chain and TCR α-chain undergo VJ recombination.
Therefore, a BCR antigen binding site that consists of both heavy and light chain (Figure 5-1) combines five gene segments that were each randomly selected from a collection of segments (Figure 5-2). This produces a tremendous range of diverse antigen binding sites and gives each lymphocyte its unique binding properties.
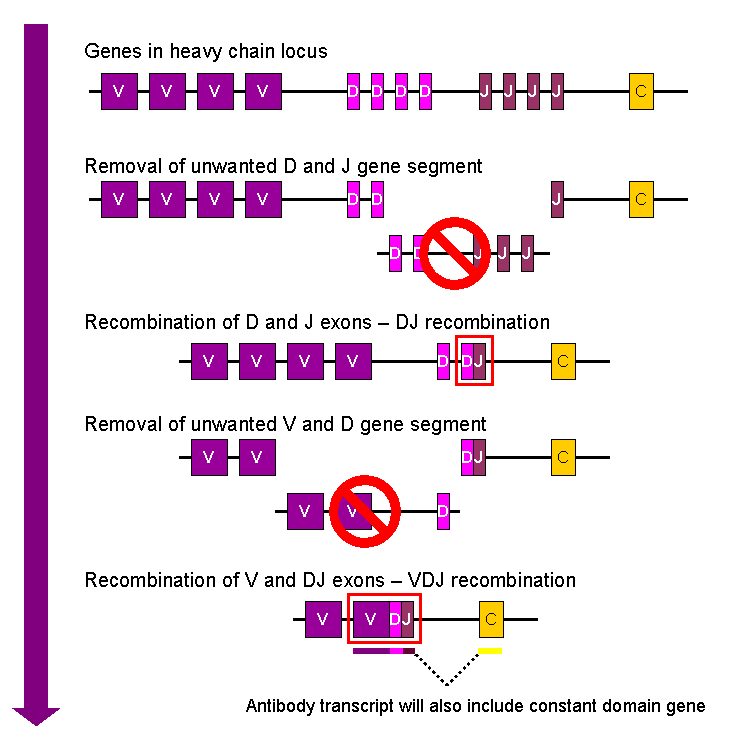
Figure 5-2 VDJ Recombination. A simplified diagram of how single variable (V), diversity (D) and Joining (J) gene segments are selected randomly from a collection and combined to achieve a tremendous diversity of antigen binding sites in B-cell and T-cell receptors. [Link to Image Description]
Image Source: By OpenStax CC BY 3.0, via Wikimedia Commons
5.2 Selection of Lymphocytes During Maturation
Lymphocytes selection is described by the clonal selection theory and involves two basic steps:
- Positive selection promotes development of functional cells and eliminates any non-functional cells by apoptosis cell death.
- Negative selection ensures that the lymphocytes do not harm human tissues by eliminating auto-reactive cells that bind human tissues with ‘self antigen’.
B-cell maturation and selection occurs in the bone marrow. First, during positive selection a checkpoint confirms that the heavy chain is functional and can bind a surrogate light chain. If this holds true the pre-B cells avoid death by apoptosis and instead proliferate rapidly. Next, negative selection involves testing the B-cell receptor, called IgM, against human antigens within the bone marrow. If B-cell clones are auto-reactive and bind human tissue, they may have an opportunity to edit their light chain. Immature B-cells that still have self-reactive receptors are retained in the bone marrow and are eventually removed in a process called clonal deletion. B-cells that survive both positive and negative clonal selection develop a second IgD receptor and are released into the bloodstream. These functional B cells migrate to lymph nodes and other secondary lymphatic tissues.
T-cells are also produced in the bone marrow but mature and undergo selection in the thymus, in a process called thymic education. T-cells enter the thymus as immature double negative (DN) thymocytes, because they lack the expression of both CD4 and CD8 co-receptors. These thymocytes initially express both CD4 and CD8 within the thymus, to become double positive (DP) thymocytes. To survive, DP thymocytes must bind to self-antigen on the surface of thymus cortical epithelial cells. After binding these T-cells receive survival signals, leading to the differentiation of either CD4+ or CD8+ single positive (SP) thymocytes.
The need for T-cells to bind self-antigen may seem counter productive to the role of these cells in detecting foreign pathogens. However, deeper within the medullary region of the thymus, the self-reactive cells are eliminated by negative selection. Negative selection involves the interaction of SP thymocytes with medullary “nurse” epithelial cells presenting a diverse range of self-antigens. SP thymocytes with high-affinity interactions to self-antigens are eliminated through apoptosis, preventing the development of T-cells that would be overly reactive against self-tissues. This process is crucial for establishing immune tolerance and preventing autoimmune responses.
Importance of leukocyte clusters of differentiation (CD) proteins
CD proteins are antigens on the surface of leukocytes that have been adopted as potent markers of cell identity and maturation. We have already alluded to the importance of these proteins when describing CD4 and CD8 as markers for T-cell subtypes. However, CD proteins can also be used more broadly to define specific immune cell populations, maturation state or even the emergence of tumor cells.
Important CD protein cell markers
- Hematopoietic stem cells give rise to all red blood cells, white blood cells and platelets. As a consequence, identification and extraction of these cells are a critical part of bone marrow transplant to recapitulate a failing immune system. The central marker for hematopoietic stem cells is CD34.
- T-cells have the common shared feature of a T-cell receptor. The CD3 protein is a defining component of the T-cell receptor. For example, helper T-cells are CD3+CD4+CD8– and cytotoxic T-cells are CD3+CD4–CD8+.
- B-cells in their naive (non-activated) form have the surface proteins CD19 and CD20.
5.3 Primary and Secondary Immune Response
Another phase of positive selection occurs when lymphocytes are exposed to their TCR-matched antigen. Following antigen presentation, helper T-cells secrete cytokines to stimulate their reproduction. Similarly, helper T-cells stimulate proliferation of B-cells, upon activation. This process of clonal expansion is a hallmark of the adaptive immune response and results in up to a thousand-fold increase in antigen-specific lymphocytes. Furthermore, some lymphocytes differentiate into memory cells that survive as a population for extended periods of time. Memory lymphocytes are immediately activated by antigen and do not require antigen presentation, making the secondary response faster. These memory lymphocytes are responsible for long-term ‘immunity’ to specific pathogens.
Consequently, the first time a pathogen antigen is encountered, the primary immune response that consists of a gradual enhancement in pathogen-specific immune defense. Every subsequent exposure to pathogen produces a secondary immune response than produces a faster and more effective immune defense.
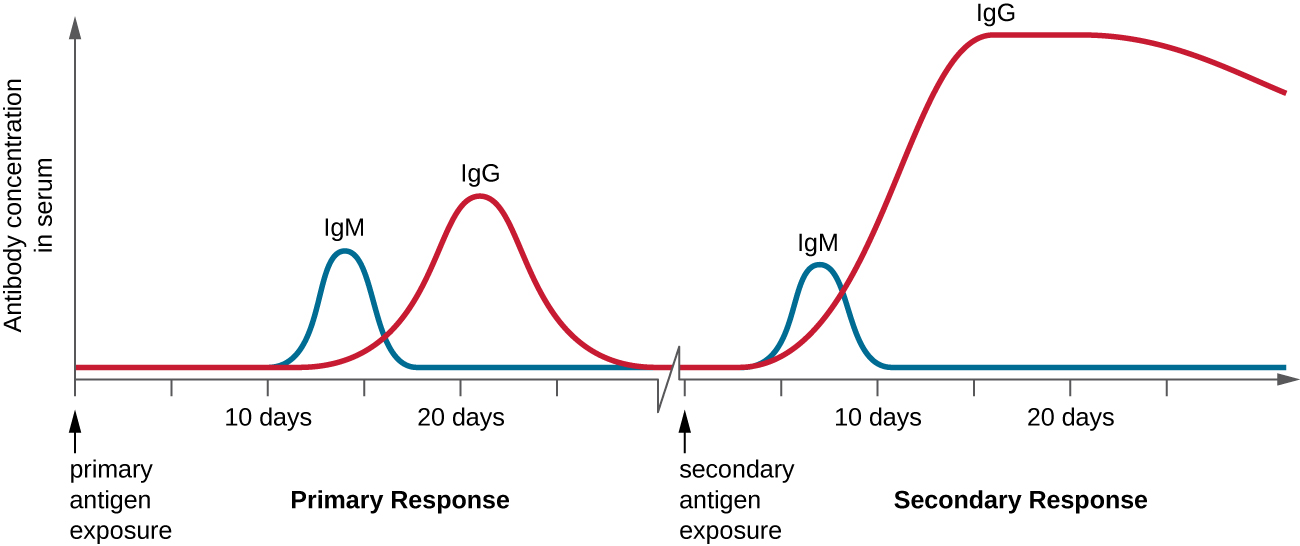
Figure 5-3 Primary and Secondary Immune Response. On first exposure to a pathogen, some antibodies (IgM and IgG) are produced. By the end of the first exposure, clonal expansion and memory lymphocytes are produced. This causes the secondary immune response to respond more quickly and to produce a far greater abundance of antibodies. [Link to Image Description]
Image Source: By OpenStax CC BY 3.0, via Wikimedia Commons
Table 5-1. Contrast of Plasma B-cells and Memory B-cells
| PLASMA B-CELLS | MEMORY B-CELLS |
| Produced is large number during infection. | Produced is small numbers following infection. |
| No surface immunoglobulin because it was secreted as antibody. | Retains surface immunoglobulin, typically IgA or IgG. |
| Short lifespan (days). | Long lifespan as the cell population is capable of self-renewal. |
| Actively secretes antibody. | Provides long term immunity through rapid response to future infection. |
5.4 Humoral Response: Antibody Production
The word humoral refers to the body fluids, where protective antibodies may be suspended to protect us from pathogens. Antibodies are B-cell receptors that are secreted into the bloodstream. Like the lymphocyte receptor, they will bind specifically only to pathogens that have the matching surface antigen molecule. Antibodies do not directly harm the target microbe. This fact is important because it means that antibodies can be used as tools to label and detect cells without harming them. Within the body, cells that are bound by antibodies are more likely to be phagocytized or bound by complement proteins that cause cell lysis.
B-Cell Activation
Naive B-cells have undergone V(D)J recombination and therefore each have an antigen-specific B-cell receptor. These naive B-cells phagocytose pathogen and debris but phagocytosis is not their main function since they only phagocytose the specific antigen, matching the B-cell receptor. Rather, the B-cell acts as an antigen presenting cell (APC) by displaying parts of the pathogen on its surface using the MHC-II complex.
Other APCs (e.g. macrophage, dendritic cell) use phagocytosis as their primary function, consuming a broad range of pathogens with surface PAMPs. These APCs present antigen similarly to a B-cell (Figure 5-4).
Both B-cells and other APCs interact with T-cells, but the outcome of these interactions are different. When a macrophage migrates from the site of infection with pathogen PAMP on its surface, it will be bound by an antigen-matched helper T-cell (Th). The Th cell receives stimulatory signals from the macrophage to become activated and can differentiate into more specialized cell subsets (see section 3.4). If the Th differentiates into the Th2 subset, these cells will specialize in B-cell activation. When a B-cell consumes and presents the same antigen to the Th2 cell, the B-cell receives signals to become activated and differentiate toward antibody secreting plasma cells as well as a less mature state representing the memory cell. In this way, cell activation is relayed from the macrophage to the Th cell and finally to the antibody-producing B-cell.
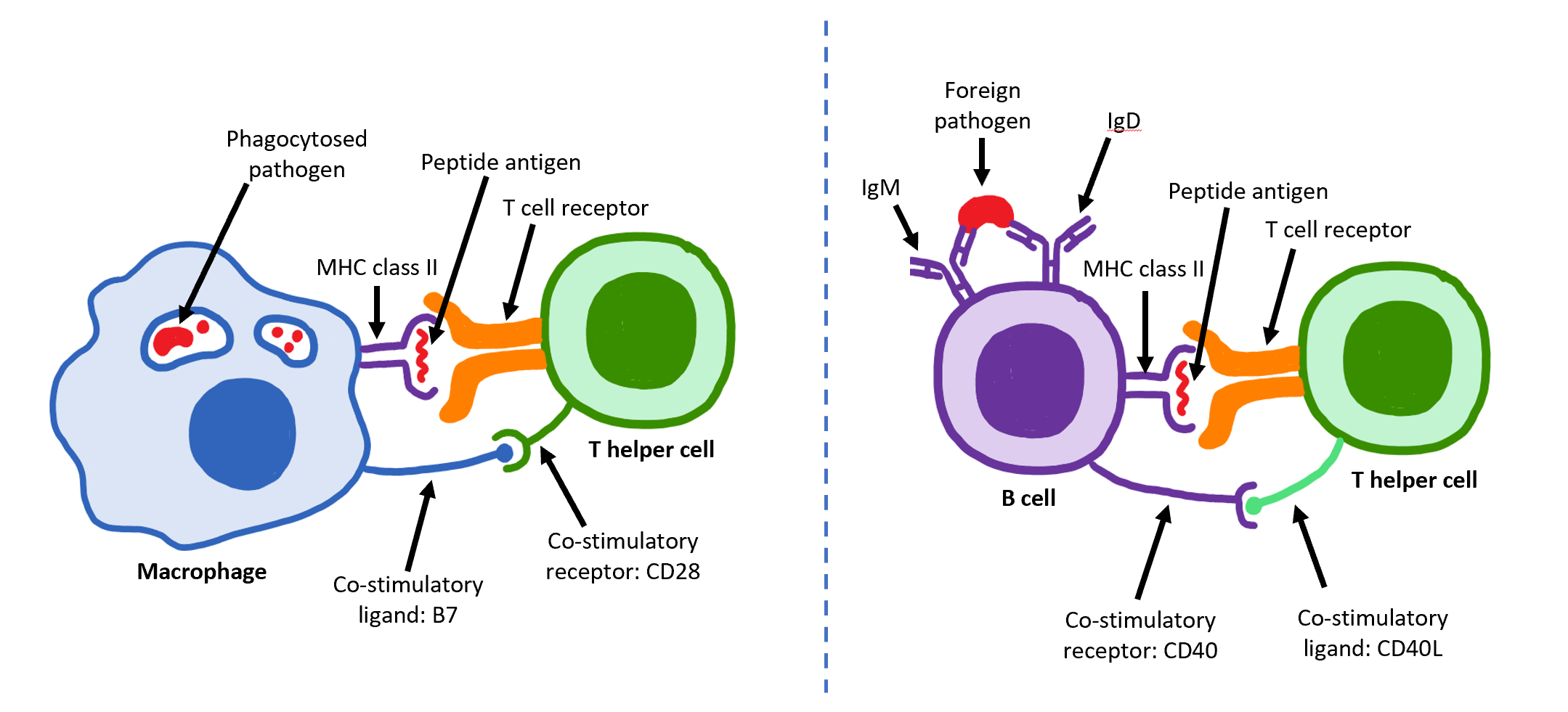
Figure 5-4 B-Cell Activation. (Left) At the site of infection, macrophages broadly consume antigen and present them via MHC-II. These cells migrate toward a local lymph node, where they interact with and activate B-cells. (Right) Within the lymph node, the B-cell phagocytosed antigen that has filtered in from the infected tissue. Upon presenting this antigen to Th2 cells, the B-cell is activated. [Link to Image Description]
Image Source: By Immcarle105 CC BY-SA 4.0, via Wikimedia Commons
T-Dependent and T-Independent B-cell Activation
The section above describes activation of B-cells upon signalling from helper T-cells in a process called T-dependent B-cell activation. However some antigens have special characteristics that allow them to activate B-cells without T-cell signalling. T-independent B-cell activation can be classified into two types:
- T-independent Type 1 (TI-1) antigens have an epitope that is bound by the B-cell receptor, as is normal for B-cells. However, other parts of the antigen bind to toll-like receptors on the surface of the B-cell and provide the additional signalling needed to activate the B-cell.
- T-independent Type 2 (TI-2) antigens have a structure where the epitope is repeated multiple times. As the B-cell bind the antigen, B-cell receptors cluster together and crosslink so they can bind the repeating epitopes. The clustering B-cell receptors provide activation signals to one another to activate the B-cell.

Figure 5-5 T-Independent B-cell Activation. TI-1 activation involves dual recognition of an antigen using a B-cell receptor (BCR) as well as a Toll-like receptor (TLR) in tandem to stimulate B-cell activation in a T-cell independent manner. TI-2 activation involves B-cell activation by an antigen with repeating units to provide a more intense activation signal and to induce T-cell independent B-cell activation. [Link to Image Description]
Image Source: By סתו כסל CC BY-SA 4.0, via Wikimedia Commons
Superantigen
A superantigen causes B-cell activation in a T-cell dependent manner but bypasses the antigen-specificity of the T-cell. Superantigens elicit a massive and uncontrolled activation of T-cells by simultaneously binding to specific regions of the TCR and MHC molecules. Staphylococcus aureus is an example of a bacterial species that has adapted superantigens as toxins to support infection. The toxin TSST-1 produces a severe dysregulated cytokine response that results in potentially fatal toxic shock syndrome.
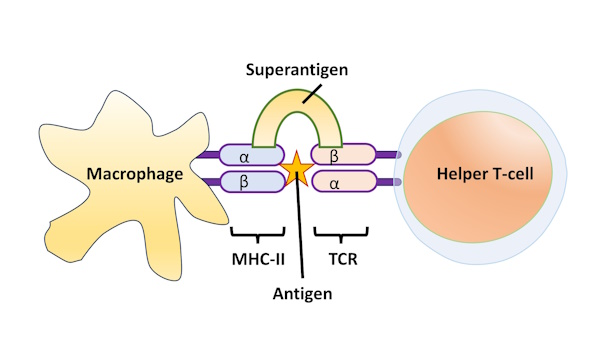
Figure 5-6 Superantigen. A macrophage presents antigen via MHC-II that is normally mismatched with the T-cell receptor (TCR). The superantigen binds the two receptors together, forcing T-cell activation despite the mis-match. [Link to Image Description]
Antibody Secretion and Class Switching
Activated B-cells differentiate into plasma cells, which secrete their surface B-cell receptor to function as antibodies, and into memory cells that persist for long periods and contribute to the secondary immune response. Plasma cells initially secrete IgM, which is secreted as a immunoglobulin monomers but assemble into pentamers that consist of five immunoglobulins connected together by a J-chain peptide.
As the primary immune response progresses, activated B-cells undergo class switching that involves recombination of the DNA encoding the immunoglobulin Fc domain. Class switching produces new immunoglobulin isotypes with distinct characteristics.
The functions of antibodies include:
Immunoprecipitation and Agglutination – Each immunoglobulin has two antigen-binding Fab regions. IgM pentamers have ten Fabs. Since each immunoglobulin can bind multiple antigen molecules, it can attach these molecules together and cause soluble molecules to become insoluble (immunoprecipitation). When the immunoglobulins connect multiple cells together, it causes the cells to clump together in the process of agglutination.
Opsonization – The Fc region of immunoglobulins are actively bound by phagocytes, making antigens and antibody-bound cells more likely to be phagocytosed.
Neutralization – Pathogens (bacteria, viruses) use proteins on their surface to adhere to and infect human tissues. By covering the surface of the pathogen, antibodies can prevent it from adhering to human tissues and infecting human cells.
Complement Fixation – The functions of antibodies are made significantly more potent by binding to complement proteins. Complement proteins form a MAC complex in the target cells and induce cell lysis. Complement proteins also enhance opsonization.
Antibody-Dependent Cell Mediated Cytotoxicity (ADCC) – The Fc region of immunoglobulins are bound by cells that may secrete toxins, such as eosinophils, macrophages and natural killer (NK) cells. This response is commonly used to eliminate pathogens that are too large to phagocytose.
| A) |
| C) |
| D) |
Figure 5-6 Effector functions of antibodies. (A) Agglutination involves attachment of antibodies to multiple cells in order to cause cells to clump together. (B) Opsonization occurs because phagocytes have receptors to the Fc region of some immunoglobulins. This causes the phagocyte to bind to and consume antibody-bound cells and material. (C) Neutralization occurs when antibodies bind the surface of cellular structures, viruses and toxins. The layer of antibodies prevent the agent from interacting with human cells, negating their harmful effects. (D) Antibody-dependent cell mediated cytotoxicity (ADCC) is similar to opsonization in that it involves leukocytes binding the Fc region of immunoglobulins. However, the target cell is typically too large to phagocytose and is therefore eliminated by secretion of toxic substances. [Link to Image Description]
Image Source: By OpenStax CC BY-SA 4.0, via OpenStax Microbiology
Figure 5-7 Characteristics of different immunoglobulin classes. [Link to Image Description]
Image Source: By OpenStax CC BY-SA 4.0, via OpenStax Microbiology
5.5 Cell Mediated Response: Overcoming Intracellular Pathogens and Cancer
In section 3.6, we observed that antigen presentation may induce helper T-cells to differentiate into Th2 cells and promote the activation of B-cells. However, helper T-cells may also differentiate into Th1 cells that secrete a different collection of cytokines. These cytokines activate macrophages and promote development of cytotoxic T-cells.
The activated macrophages will be induced to phagocytose pathogen more efficiently and will secrete cytokines that promote inflammation at the site of infection. For this reason, the Th1 response is regarded as a pro-inflammatory immune response.
The cytotoxic T-cells have CD8 on their surface, which causes them to attach to virtually all cells via binding MHC-I. Normal human cells routinely load peptides from inside the cell onto MHC-I and display these peptides on the cell surface. If a cell becomes infected or produces abnormal cancer proteins, those foreign proteins would also be displayed by MHC-I. The T-cell receptor then scans the MHC-I and peptides on the cell surface and, if an abnormal peptide is recognized, the cytotoxic T-cell will kill the target cell. Even though MHC-I displays normal human peptide as well as any viral or cancer peptide, cytotoxic T-cells will not respond to the normal peptides because of negative selection during maturation.
The binding of a T-cell to its target depends on an immune synapse
When a T-cell encounters an antigen-presenting cell (APC), one factor that decides whether the two cells will bind is compatibility with MHC type. Recall, CD8 binds MHC-I and CD4 binds MHC-II. However, a sustained immune synapse that results in T-cell activation requires an orchestrated interplay of molecular interactions between receptors on the T-cell and the APC. The central supramolecular activation cluster (cSMAC) forms, concentrating T-cell receptor (TCR)-MHC interactions, while the peripheral SMAC (pSMAC) and distal SMAC (dSMAC) arrange adhesion and signaling molecules. This dynamic structure facilitates the transmission of signals from the TCR complex to the T-cell’s interior, initiating a cascade of events that lead to T-cell activation, proliferation, and subsequent immune responses.
The cytotoxic T-cell uses various methods to kill the target cell:
Perforins are related to complement proteins and are capable of producing pores in the target cell membrane.
Granzymes are enzymes that enter through the pores and induce apoptosis death in the target cell.
Fas ligand (FasL) is a protein on the surface of cytotoxic T-cells. FasL binds the Fas receptor on the target cell and induces an apoptosis suicide pathway in that cell.
Figure 5-8 Effector function of cytotoxic T-cell. Human cells display their internal peptides using MHC-I. This means that an infected cell will also present the peptides from that intracellular pathogen. Cytotoxic T-cells are characterized by CD8 on their surface as well as an antigen-specific T-cell receptor. CD8 will attach the cytotoxic T-cell to MHC-I, while the T-cell receptor recognizes the pathogen peptide. Upon recognizing the infected cell, the cytotoxic T-cell will destroy the cell with granzymes and perforins. [Link to Image Description]
5.6 Immune Tolerance
To function properly, the immune system must be able to recognize and respond to foreign invaders, such as pathogens, while avoiding inappropriate responses against the body’s own tissues or microbiome that is a normal component of the body surface. Immune tolerance refers to the collective mechanisms by which the immune system prevents undesirable immune responses to self-antigens and commensal microbes.
Immune tolerance can be categorized into two types:
Central tolerance involves the negative selection process that occurs during the development of immune cells in the thymus (T cells) and bone marrow (B cells). Developing lymphocytes that recognize self-antigens with high affinity are eliminated or rendered non-functional. This ensures that only lymphocytes with the ability to recognize foreign antigens are allowed to mature and participate in immune responses.
Peripheral Tolerance involves suppression or elimination of self-reactive lymphocytes that are not eliminated by central tolerance. The mechanisms for peripheral tolerance include:
- Regulatory T cells (Tregs): Like helper T-cells, Treg respond to specific antigen. However, they suppress the activity of other immune cells either through direct interaction with these cells or by secretion of immunosuppressive cytokines.
- Clonal Anergy and Clonal Deletion: Anergy and deletion may occur in both B-cells and T-cells but their role in peripheral tolerance is characteristic of T-cells. Antigen presentation normally occurs in the context of inflammation, where the antigen presenting cell presents antigen via MHC-II but also expresses proteins called co-stimulatory proteins. If a T-cell interacts with antigen outside of the context of infection and inflammation, the T-cell clones may be deleted by apoptosis or may expand into a non-responsive cell population (anergy).
- Immune Privilege: Certain tissues or organs in the body, such as the eyes, brain, and reproductive organs have specialized mechanisms that limit immune responses within their boundaries, reducing the likelihood of immune-mediated damage to these critical structures.
Summary
- B-cells and T-cells are lymphocytes that are involved in the immune response. B-cells secrete antibodies, while T-cells may induce an immune response (Helper T), suppress an immune response (Regulatory T) or kill virus-infected or cancer cells (Cytotoxic T).
- Each B-cell and T-cell has a unique receptor that is produced by randomly assorting genomic DNA segments, as V(D)J recombination. This process defines the antigen to which the lymphocyte will react.
- Lymphocytes undergo positive selection to ensure they are functional as well as negative selection that eliminates autoreactive cells.
- In addition to the ‘effector’ lymphocytes, B-cell and T-cell memory cells produce immunological memory that generates a faster and stronger immune response to future infections.
- B-cell activation requires that the B-cell binds its antigen via the B-cell receptor, phagocytoses and presents the antigen using MHC-II. Independently, an antigen presenting cell (APC) also phagocytizes and presents the same antigen. A helper T-cell must be activated by the APC and then interact with the antigen-bearing B-cell in order to secrete the cytokines needed to activate that B-cell.
- B-cells can be activated independently of helper T-cells by additional toll-like receptor signalling (TI-1) or by repeating epitopes (TI-2).
- T-cells can be activated independently of a matched antigen via superantigen
- B-cells secrete antibodies that have five basic functions: (1) Agglutination/Immunoprecipitation, (2) Opsonization, (3) Neutralization or (4) Complement fixation, (5) Antibody-dependent cell-mediated cytotoxicity
- After B-cell activation, some cells undergo class switching. Each antibody class has different roles in the immune response.
- Cytotoxic T-cells kill virus-infected or cancer cells by lysing (“bursting”) the cell using perforin proteins, or by inducing apoptotic death in that cell using granzymes or Fas ligand signalling
- Not all lymphocytes specifically induce immune response. Tolerance of self-cells or commonly present microbes may be achieved by elimination of some lymphocyte clones as well as regulatory T-cells to actively suppress the immune response.
Chapter Review
Case Study Review
specialized epithelial tissues that secrete mucus.
Pathogen associated molecular patterns (PAMPs) are the general features that our innate immune system uses to identify foreign agents, like bacteria and viruses. PAMPs are different from "antigens", which are molecules that are specifically identified by cells and antibodies of the adaptive immune system.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}