1203 Chapter 28. Development and Inheritance
28.1 Fertilization
Learning Objectives
By the end of this section, you will be able to:
- Describe the process of fertilization, specifying the site where it normally occurs, the general events that occur during fertilization, and the product of fertilization
- Explain the phenomenon of multiple births in humans
Fertilization occurs when a sperm and an oocyte (egg) combine and their nuclei fuse. Because each of these reproductive cells is a haploid cell containing half of the genetic material needed to form a human being, their combination forms a diploid cell. This new single cell, called a zygote, contains all of the genetic material needed to form a human—half from the mother and half from the father.
Transit of Sperm
Fertilization is a numbers game. During ejaculation, hundreds of millions of sperm (spermatozoa) are released into the vagina. Almost immediately, millions of these sperm are overcome by the acidity of the vagina (approximately pH 3.8), and millions more may be blocked from entering the uterus by thick cervical mucus. Of those that do enter, thousands are destroyed by phagocytic uterine leukocytes. Thus, the race into the uterine tubes, which is the most typical site for sperm to encounter the oocyte, is reduced to a few thousand contenders. Their journey—thought to be facilitated by uterine contractions—usually takes from 30 minutes to 2 hours. If the sperm do not encounter an oocyte immediately, they can survive in the uterine tubes for another 3–5 days. Thus, fertilization can still occur if intercourse takes place a few days before ovulation. In comparison, an oocyte can survive independently for only approximately 24 hours following ovulation. Intercourse more than a day after ovulation will therefore usually not result in fertilization.
During the journey, fluids in the female reproductive tract prepare the sperm for fertilization through a process called capacitation, or priming. The fluids improve the motility of the spermatozoa. They also deplete cholesterol molecules embedded in the membrane of the head of the sperm, thinning the membrane in such a way that will help facilitate the release of the lysosomal (digestive) enzymes needed for the sperm to penetrate the oocyte’s exterior once contact is made. Sperm must undergo the process of capacitation in order to have the “capacity” to fertilize an oocyte. If they reach the oocyte before capacitation is complete, they will be unable to penetrate the oocyte’s thick outer layer of cells.
Contact Between Sperm and Oocyte
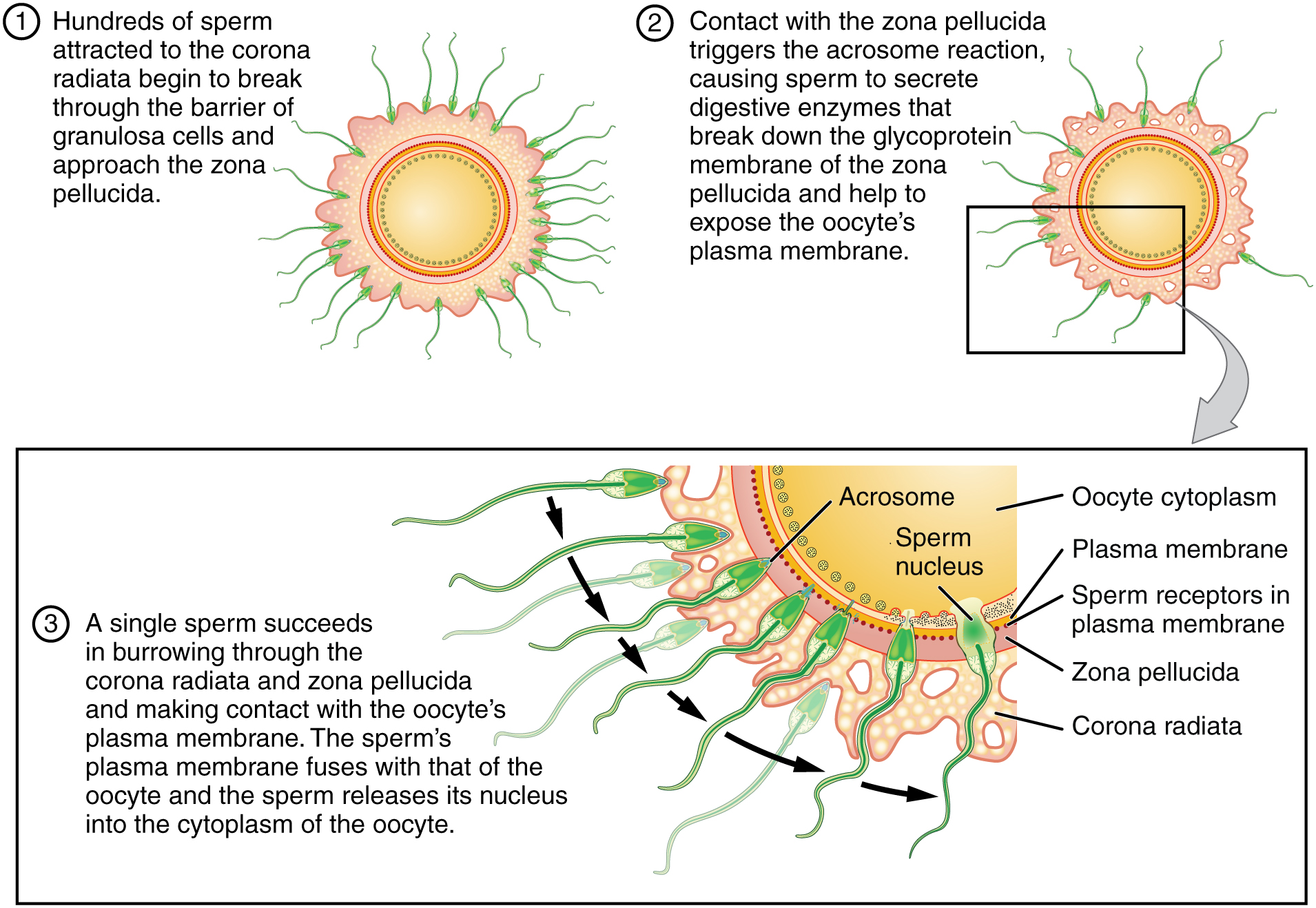
Upon ovulation, the oocyte released by the ovary is swept into—and along—the uterine tube. Fertilization must occur in the distal uterine tube because an unfertilized oocyte cannot survive the 72-hour journey to the uterus. As you will recall from your study of the oogenesis, this oocyte (specifically a secondary oocyte) is surrounded by two protective layers. The corona radiata is an outer layer of follicular (granulosa) cells that form around a developing oocyte in the ovary and remain with it upon ovulation. The underlying zona pellucida (pellucid = “transparent”) is a transparent, but thick, glycoprotein membrane that surrounds the cell’s plasma membrane.
As it is swept along the distal uterine tube, the oocyte encounters the surviving capacitated sperm, which stream toward it in response to chemical attractants released by the cells of the corona radiata. To reach the oocyte itself, the sperm must penetrate the two protective layers. The sperm first burrow through the cells of the corona radiata. Then, upon contact with the zona pellucida, the sperm bind to receptors in the zona pellucida. This initiates a process called the acrosomal reaction in which the enzyme-filled “cap” of the sperm, called the acrosome, releases its stored digestive enzymes. These enzymes clear a path through the zona pellucida that allows sperm to reach the oocyte. Finally, a single sperm makes contact with sperm-binding receptors on the oocyte’s plasma membrane (Figure 1). The plasma membrane of that sperm then fuses with the oocyte’s plasma membrane, and the head and mid-piece of the “winning” sperm enter the oocyte interior.
How do sperm penetrate the corona radiata? Some sperm undergo a spontaneous acrosomal reaction, which is an acrosomal reaction not triggered by contact with the zona pellucida. The digestive enzymes released by this reaction digest the extracellular matrix of the corona radiata. As you can see, the first sperm to reach the oocyte is never the one to fertilize it. Rather, hundreds of sperm cells must undergo the acrosomal reaction, each helping to degrade the corona radiata and zona pellucida until a path is created to allow one sperm to contact and fuse with the plasma membrane of the oocyte. If you consider the loss of millions of sperm between entry into the vagina and degradation of the zona pellucida, you can understand why a low sperm count can cause male infertility.
When the first sperm fuses with the oocyte, the oocyte deploys two mechanisms to prevent polyspermy, which is penetration by more than one sperm. This is critical because if more than one sperm were to fertilize the oocyte, the resulting zygote would be a triploid organism with three sets of chromosomes. This is incompatible with life.
The first mechanism is the fast block, which involves a near instantaneous change in sodium ion permeability upon binding of the first sperm, depolarizing the oocyte plasma membrane and preventing the fusion of additional sperm cells. The fast block sets in almost immediately and lasts for about a minute, during which time an influx of calcium ions following sperm penetration triggers the second mechanism, the slow block. In this process, referred to as the cortical reaction, cortical granules sitting immediately below the oocyte plasma membrane fuse with the membrane and release zonal inhibiting proteins and mucopolysaccharides into the space between the plasma membrane and the zona pellucida. Zonal inhibiting proteins cause the release of any other attached sperm and destroy the oocyte’s sperm receptors, thus preventing any more sperm from binding. The mucopolysaccharides then coat the nascent zygote in an impenetrable barrier that, together with hardened zona pellucida, is called a fertilization membrane.
The Zygote
Recall that at the point of fertilization, the oocyte has not yet completed meiosis; all secondary oocytes remain arrested in metaphase of meiosis II until fertilization. Only upon fertilization does the oocyte complete meiosis. The unneeded complement of genetic material that results is stored in a second polar body that is eventually ejected. At this moment, the oocyte has become an ovum, the female haploid gamete. The two haploid nuclei derived from the sperm and oocyte and contained within the egg are referred to as pronuclei. They decondense, expand, and replicate their DNA in preparation for mitosis. The pronuclei then migrate toward each other, their nuclear envelopes disintegrate, and the male- and female-derived genetic material intermingles. This step completes the process of fertilization and results in a single-celled diploid zygote with all the genetic instructions it needs to develop into a human.
Most of the time, a woman releases a single egg during an ovulation cycle. However, in approximately 1 percent of ovulation cycles, two eggs are released and both are fertilized. Two zygotes form, implant, and develop, resulting in the birth of dizygotic (or fraternal) twins. Because dizygotic twins develop from two eggs fertilized by two sperm, they are no more identical than siblings born at different times.
Much less commonly, a zygote can divide into two separate offspring during early development. This results in the birth of monozygotic (or identical) twins. Although the zygote can split as early as the two-cell stage, splitting occurs most commonly during the early blastocyst stage, with roughly 70–100 cells present. These two scenarios are distinct from each other, in that the twin embryos that separated at the two-cell stage will have individual placentas, whereas twin embryos that form from separation at the blastocyst stage will share a placenta and a chorionic cavity.

