56 EWE book testing – Villy Christensen
[latexpage]
Source: https://pressbooks.bccampus.ca/ewemodel/chapter/introduction-to-ecospace/
About Ecospace
The Ecospace model is a spatially explicit time dynamic model based on the Ecopath mass-balance and Ecosim time dynamic routines [1] [2]. It applies the same set of differential equations as used in Ecosim, executed for each cell in a grid of cells. In Ecosim, a set of differential equations is defined based on the biomass change during time for consumer functional groups, expressed as
[latex]\begin{equation}\frac{dB_i}{dt}=g_i\cdot\sum\limits_{i=1}^{n}Q_{ij}-\sum\limits_{j=1}^{n}Q_{ji}+I_i-(F_{it}+e_i+M0_{it})\cdot B_{it}\tag{}\label{one}\end{equation}[/latex]
where Bit is the biomass of i at time t, gi is the growth efficiency, Ii is the immigration rate; Fit is the mortality rate due to harvesting (fishing mortality); ei is the emigration rate; and M0i the other mortality (mortality not explained in the model). The terms Qij and Qji represent the consumption due to predation by i on j, and by j on i, respectively. For primary producers, the term f (Bit)) represents the growth term as function of the group biomass [3].
The consumption rates Qij are based on the foraging arena theory (see chapter), where the biomass of prey i is split between a vulnerable (Vij) and a non-vulnerable (Bi-Vij) component. The transfer rate, called vulnerability (υij) between the two fractions determines the vulnerable biomass at time interval dt:
[latex]\frac{dV_{ij}}{dt}=v_{ij}\cdot(B_i-V_{ij})-v_{ij}\cdot V_{ij}-\frac{a_{ij}V_{ij}B_j}{1+h_ja_{ij}V_{ij}}\label{two})[/latex]
where aij is the effective search rate for the predator j, and hj is handling time for the predator. The vulnerability parameter υij is a function of the maximum increase in predation mortality under the given predator/prey conditions (see vulnerability multiplier chapter). High values of υij imply large proportions of biomass (Bi) vulnerable to predator j (Vij), and thus imply Vij = Bi, and that the predator j is far from its carrying capacity with regards to prey i.
In Ecospace, the spatial extent of the ecosystem is represented by a grid of cells, each of which can be defined as land or water and, and have a habitat type assigned to the cell. Ecospace represents the biomass (B) and consumption (Q) dynamics over a two-dimensional space as well as time [4]. Space, time, and state are considered discrete variables by using the Eulerian approach, which treats movements as ‘flows’ of organisms among fixed spatial reference cells.
In the original Ecospace model [5], a first step of parameterizing entails the definition of a basemap based on habitat information (depth strata, bottom type, etc.) in the study area. Species preferences are then assigned to these habitat types based on the biology and ecology of the species included in each functional group of the ecosystem model, their depth distributions, their preferred sediment type, etc. In addition, the original Ecospace model required for habitat definitions,
- the dispersion rate of each functional group in ‘preferred’ habitats,
- the relative dispersal rate in ‘non-preferred’ habitats, and
- the relative feeding rate in non-preferred habitat by functional group.
Fishing fleets can be depicted as operating in a specific region and cells can be defined as protected areas.
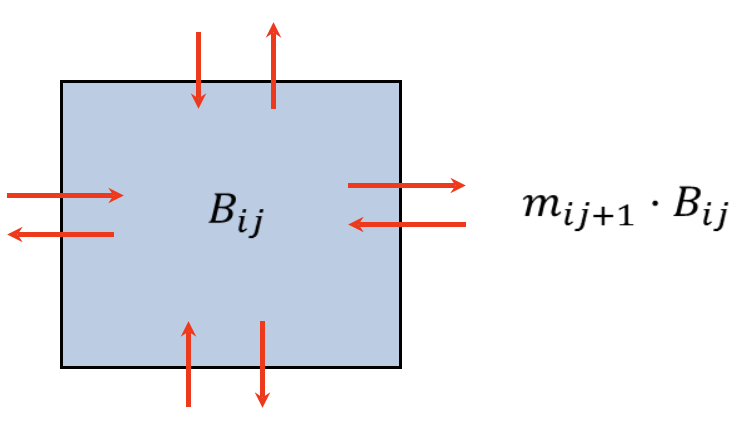
Figure 1. For each cell, the inbound dispersal rate Ii is the sum of emigration flows from the four surrounding cells, while the outbound instantaneous dispersal rates mi from a given cell in Ecospace vary based on the pool type, cell conditions/habitat, and response of organisms to predation risk and feeding conditions
Moreover, relative variations of primary productivity and fishing costs can be defined for the initial conditions of the model. For trophic interactions, fishing, and movement calculations, biomass is considered as homogeneous within each cell and movement of biomass and flows is allowed across the borders to adjacent cells. For each cell, the immigration rate Ii of Eq. \ref{two} is assumed to consist of up to four emigration flows from the surrounding cells (Figure 1). The emigration flows (Bout,rci) are in turn similarly represented as instantaneous movement rates mi times the biomass density in the cell (Brci):
[latex]B_{out,rci}=\sum\limits_{d=1}^{4}m_{id}\cdot B_{rci} \label{three}[/latex]
where (rci) represents cell row and column for group i, and d is movement direction (up, down, left or right).
The instantaneous emigration rates mi from a given cell in Ecospace are assumed to vary based on the functional group, habitat preferences, and responses of organisms to depredation risk and feeding conditions. The probability of movement of organisms towards favourable habitats was in the original Ecospace formulation calculated by means of a ‘habitat gradient function’ for each mapped habitat type and species or group i. Biomass dynamics in unsuitable cells were modified by predicting higher rates of emigration, lower feeding rates, and/or higher vulnerability to predation, a complex gradient calculation to modify dispersal rates is used to direct biomass toward suitable cells.
In more recent versions of Ecospace, a habitat capacity model has been included to estimate cell-specific continuous habitat suitability factor, where the area that species can feed in each cell is determined by functional responses to multiple environmental factors. See the habitat capacity chapter. It is optional whether to use a habitat and/or habitat suitability for any given group, though in many recent applications habitat suitability is used predominantly while habitats mainly are used for defining where fleets can operate.
Attribution
The first section of this chapter is based on de Mutsert K, Marta Coll, Jeroen Steenbeek, Cameron Ainsworth, Joe Buszowski, David Chagaris, Villy Christensen, Sheila J.J. Heymans, Kristy A. Lewis, Simone Libralato, Greig Oldford, Chiara Piroddi, Giovanni Romagnoni, Natalia Serpetti, Michael Spence, Carl Walters. 2023. Advances in spatial-temporal coastal and marine ecosystem modeling using Ecopath with Ecosim and Ecospace. Treatise on Estuarine and Coastal Science, 2nd Edition. Elsevier. https://doi.org/10.1016/B978-0-323-90798-9.00035-4, adapted with permission, License Number 5651431253138.
The second section of the chapter is partly based on Christensen, V, M Coll, J Steenbeek, J Buszowski, D Chagaris, and CJ Walters. 2014. Representing variable habitat quality in a spatial food web model. Ecosystems 17(8): 1397-1412. https://doi.org/10.1007/s10021-014-9803-3.
Rather than citing this chapter, please cite the sources
- Walters C, Christensen V, Pauly D. 1997. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Reviews in Fish Biology and Fisheries 7: 139-172. https://doi.org/10.1023/A:1018479526149 ↵
- Christensen V, Walters C. 2004. Ecopath with Ecosim: methods, capabilities and limitations. Ecological Modelling 72: 109-139. https://doi.org/10.1016/j.ecolmodel.2003.09.003 ↵
- Christensen and Walters. 2004. op. cit. ↵
- Walters C, Pauly D, Christensen V. 1999. Ecospace: prediction of mesoscale spatial patterns in trophic relationships of exploited ecosystems, with emphasis on the impacts of marine protected areas. Ecosystems 2: 539-554. https://doi.org/10.1007/s100219900101 ↵
- Walters et al. 1999. op. cit. ↵
