50 Introduction to Ecospace
Setting the stage
Marine ecosystems are complex systems affected by the state of the environment and a myriad of human activities. The full impact of these human activities on the complex ecosystem are becoming more critical to understand, and are now requested by agencies concerned with numerous policies and strategies that involve spatial management actions (in Europe, for instance by the Common Fisheries Policy, Marine Strategy Framework Directive, Farm to Fork, Zero Pollution and Biodiversity Strategies). All of these have divergent end-points, but all require an understanding of how multiple forms of human activities impact marine ecosystems. To better manage our impact on marine ecosystems, and notably consider trade-offs in management, there is a need to advance scientific capabilities to provide both quantitative descriptions and quantitative evaluations of the effect of spatial management interventions, factoring in plausible future changes in climate and human activities.
Spatial-temporal ecosystem modeling has grown in capacity, complexity, and focus to undertake such complex tasks, and is increasingly considered an indispensable tool to contribute to policy and management, including multi-sectoral Ecosystem Based Management (EBM) and Marine Spatial Planning (MSP).
Among the tools available, the temporal and spatial dynamic model EwE approach is of special interest for its wide range of applications across ecosystem types. Ecospace has notably been used to contribute to EBM, most often to a subset of EBM, Ecosystem-Based Fisheries Management, or EBFM. Applications of Ecospace include the evaluation of spatial trophic interaction patterns, modeling of species distribution based on habitat suitability, the assessment of Marine Protected Area (MPA) placement and connectivity, harvest allocations and, more recently, environmental impact analysis and the assessment of episodic mortality events, effects of changes in nutrient inputs, climate change, and cumulative impacts (e.g.,[1] [2] [3] [4] [5] [6] [7] [8] [9] [10] [11] [12] [13] [14] [15] [16] [17] [18].
The development of Ecospace has involved an evolutionary process where many additional capabilities have been developed over the years in response to requests by users. The more recent advancements include the Habitat Foraging Capacity Model (HFC[19])and the Spatial Temporal Data Framework (STDF[20]), which have enabled Ecospace applications to fully consider climate variability and change, taking into account different types of uncertainty[21] [22]. These innovations and a substantial increase in the various applications of EwE[23], provided a call for an update of the Ecospace, which led to a book chapter by de Mutsert et al.[24] upon which a number of the chapters in the present text book are based (as attributed). The present text book and the EwE User Guide seek to present the most up-to-date guidelines for using and understanding Ecospace’s capabilities and challenges.
About Ecospace
The Ecospace model is a spatially explicit time dynamic model based on the Ecopath mass-balance and Ecosim time dynamic routines[25] [26]. It applies the same set of differential equations as used in Ecosim, executed for each functional group and cell in a grid of cells. In Ecosim, a set of differential equations is defined based on the biomass components of change for consumer functional groups, expressed as
[latex]\frac{dB_i}{dt}=g_i\cdot\sum\limits_{j=1}^{n}Q_{ji}-\sum\limits_{j=1}^{n}Q_{ij}+I_i-(F_{it}+e_i+M0_{it})\cdot B_{it}\tag{1}[/latex]
where Bit is the biomass of i at time t, gi is the growth efficiency, Ii is the immigration rate; Fit is the mortality rate due to harvesting (fishing mortality); ei is the emigration rate; and M0i the other mortality (mortality not explained in the model). The terms Qji and Qij represent the consumption due to predation by j on i, and by i on j, respectively. For primary producers, the consumption rate term is replaced by a production rate function f (Bit)) represents primary production rate as a function of the group biomass[27]; that function is nonlinear, representing competition effects for light and nutrients.
The consumption rates Qji are predicted using equations from foraging arena theory (see chapter), where the biomass of prey i is split between a vulnerable (Vij) and a non-vulnerable (Bi-Vij) component. The vulnerability exchange parameters used in predicting the various Qji mainly represent spatial distribution and movement behaviors at very fine scales, typically far smaller than the size of Ecospace spatial grid cells.
In Ecospace, the spatial extent of the ecosystem is represented by a grid of cells, each of which can be defined as land or water, and each cell can have characteristics or attributes like a habitat type. Ecospace then represents the biomass (B) and consumption (Q) dynamics over a two-dimensional space as well as time[28]. Space, time, and state are considered discrete variables by using the Eulerian approach, which treats movements as "flow" of organisms among fixed spatial reference cells.
In the original Ecospace model[29], a first step of parameterizing entailed the definition of a base map based on habitat information (depth strata, bottom type, etc.) in the study area. Species preferences were then (and still can be if more elaborate spatio-temporal habitat use functions are not used) assigned to these habitat types based on the biology and ecology of the species included in each functional group of the ecosystem model, their depth distributions, their preferred sediment type, etc. In addition, the original Ecospace model required for habitat definitions,
- the dispersal (spatial mixing) rate of each functional group in "preferred" habitats,
- the relative dispersal rate in "non-preferred" habitats, and
- the relative feeding rate in non-preferred habitat by functional group.
Fishing mortality rate for each cell can represent effects of fishing effort by multiple fishing fleets, and each fishing fleet can be depicted as operating in a specific region and habitat type, and cells can be defined as protected areas for particular or all fishing fleets. Fishing effort is assumed to move between grid cells over time in response to spatial and temporal variation in profitability of fishing.
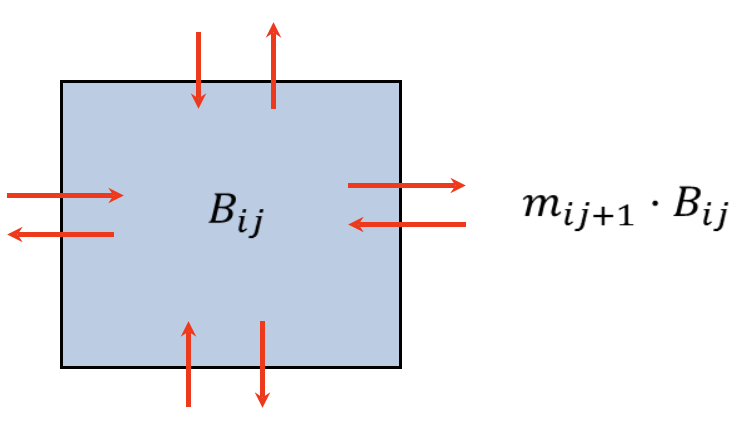
Figure 1. For each cell, the inbound dispersal rate Ii is the sum of emigration flows from the four surrounding cells, while the outbound instantaneous dispersal rates mi from a given cell in Ecospace make up the basic Ecosim emigration rates ei, and vary based on the pool type, cell conditions/habitat, and response of organisms to predation risk and feeding conditions.
Moreover, spatial variations of primary productivity and fishing costs can be defined as initial conditions for the basic model.
For trophic interactions, fishing, and movement calculations, biomass is considered as homogeneous within each cell and movement of biomass and flows is allowed across the borders to adjacent cells. For each cell, the immigration rate Ii of Eq. 1 is assumed to consist of up to four emigration flows from the surrounding cells (Figure 1). The emigration flows (Bout,rci) are in turn similarly represented by instantaneous movement rates mi times the biomass density in the cell (Brci) with the sum of those loss rates like mi,j+1 representing the emigration rate ei in eq. 1:
[latex]B_{out,rci}=\sum\limits_{d=1}^{4}m_{id}\cdot B_{rci} \tag{3}[/latex]
where (rci) represents cell row and column for group i, and d is movement direction (up, down, left or right).
The instantaneous emigration rates mi,d from a given cell in Ecospace are assumed to vary based on the functional group, habitat preferences, and can be set to vary with trophic conditions within each cell (to model responses of organisms to predation risk and feeding conditions). The probability of movement of organisms towards favourable habitats was in the original Ecospace formulation calculated by means of a "habitat gradient function" for each mapped habitat type and species or group i. Biomass dynamics in unsuitable cells were modified by predicting higher rates of emigration, lower feeding rates, and/or higher vulnerability to predation, and a complex gradient calculation continues to be used so as to modify dispersal rates to cause higher movement rates of biomass toward suitable cells.
In more recent versions of Ecospace, a habitat capacity model has been included to estimate cell-specific continuous habitat suitability factors where the area that species can feed in each cell is determined by functional responses to multiple environmental factors[30]. See the habitat capacity chapter. It is optional whether to use a habitat and/or habitat suitability for any given group, though in many recent applications habitat suitability is used predominantly while habitats mainly are used for defining where fleets can operate.
The very large equation system represented by eq. 1 with mixing terms is solved numerically on one month time steps using an implicit integration method (BDF2, second order backward differentiation), that works very well for long-lived species but tends to predict short lived species to change more slowly than Ecosim predicts. As in Ecosim, the Ecospace BDF integration does not "see" the very rapid boom-bust dynamics that can be exhibited by groups with high P/B (e.g. phytoplankton and short-lived zooplankters), but instead predicts an average biomass for these groups.
For advanced applications involving multi-stanza groups, there are two Ecospace option[31]. The first "multi-stanza" solution option. is to keep track of overall multi-stanza numbers at age over the map while predicting local variation in abundance from concentration patterns predicted from spatial mij variations of the differential equation system. The second or "IBM" option is to divide the multi-stanza recruitment numbers at each time step into a large number of packets of individuals, then simulate random and directed movements of these packets over the map as the organisms grow. Details of the IBM equations are presented later in the chapter on Spatial implementation of multi-stanza and IBM.
Quiz
Attribution The first section of this chapter is based on de Mutsert K, Marta Coll, Jeroen Steenbeek, Cameron Ainsworth, Joe Buszowski, David Chagaris, Villy Christensen, Sheila J.J. Heymans, Kristy A. Lewis, Simone Libralato, Greig Oldford, Chiara Piroddi, Giovanni Romagnoni, Natalia Serpetti, Michael Spence, Carl Walters. 2023. Advances in spatial-temporal coastal and marine ecosystem modeling using Ecopath with Ecosim and Ecospace. Treatise on Estuarine and Coastal Science, 2nd Edition. Elsevier. https://doi.org/10.1016/B978-0-323-90798-9.00035-4, adapted with permission, License Number 5651431253138.
The second section of the chapter is partly based on Christensen, V, M Coll, J Steenbeek, J Buszowski, D Chagaris, and CJ Walters. 2014. Representing variable habitat quality in a spatial food web model. Ecosystems 17(8): 1397-1412. https://doi.org/10.1007/s10021-014-9803-3.
Rather than citing this chapter, please cite the sources
- Alexander, K.A., Meyjes, S.A., Heymans, J.J., 2016. Spatial ecosystem modelling of marine renewable energy installations: Gauging the utility of Ecospace. Ecological Modelling, Ecopath 30 years – Modelling ecosystem dynamics: beyond boundaries with EwE 331, 115–128. https://doi.org/10.1016/j.ecolmodel.2016.01.016 ↵
- Coll, M., Pennino, M.G., Steenbeek, J., Sole, J., Bellido, J.M., 2019. Predicting marine species distributions: Complementarity of food-web and Bayesian hierarchical modelling approaches. Ecological Modelling 405, 86–101. https://doi.org/10.1016/j.ecolmodel.2019.05.005 ↵
- Dahood, A., de Mutsert, K., Watters, G.M., 2020. Evaluating Antarctic marine protected area scenarios using a dynamic food web model. Biological Conservation 251, 108766. https://doi.org/10.1016/j.biocon.2020.108766 ↵
- De Mutsert, K., Lewis, K., Milroy, S., Buszowski, J., Steenbeek, J., 2017a. Using ecosystem modeling to evaluate trade-offs in coastal management: Effects of large-scale river diversions on fish and fisheries. Ecological Modelling 360, 14–26. https://doi.org/10.1016/j.ecolmodel.2017.06.029 ↵
- De Mutsert, K., Lewis, K.A., White, E.D., Buszowski, J., 2021. End-to-End Modeling Reveals Species-Specific Effects of Large-Scale Coastal Restoration on Living Resources Facing Climate Change. Front. Mar. Sci. 8. https://doi.org/10.3389/fmars.2021.624532 ↵
- Espinosa-Romero, M.J., Gregr, E.J., Walters, C., Christensen, V., Chan, K.M.A., 2011. Representing mediating effects and species reintroductions in Ecopath with Ecosim. Ecological Modelling 222, 1569–1579. https://doi.org/10.1016/j.ecolmodel.2011.02.008 ↵
- Fouzai, N., Coll, M., Palomera, I., Santojanni, A., Arneri, E., Christensen, V., 2012. Fishing management scenarios to rebuild exploited resources and ecosystems of the Northern-Central Adriatic (Mediterranean Sea). Journal of Marine Systems 102–104, 39–51. https://doi.org/10.1016/j.jmarsys.2012.05.003 ↵
- Libralato, S., Solidoro, C., 2009. Bridging biogeochemical and food web models for an End-to-End representation of marine ecosystem dynamics: The Venice lagoon case study. Ecological Modelling 220, 2960–2971. https://doi.org/10.1016/j.ecolmodel.2009.08.017 ↵
- Martell, S.J.D., Essington, T.E., Lessard, B., Kitchell, J.F., Walters, C.J., Boggs, C.H., 2005. Interactions of productivity, predation risk, and fishing effort in the efficacy of marine protected areas for the central Pacific. Can. J. Fish. Aquat. Sci. 62, 1320–1336. https://doi.org/10.1139/f05-114 ↵
- Masi, M.D., Ainsworth, C.H., Kaplan, I.C., Schirripa, M.J., 2018. Interspecific Interactions May Influence Reef Fish Management Strategies in the Gulf of Mexico. Mar Coast Fish 10, 24–39. https://doi.org/10.1002/mcf2.10001 ↵
- Okey, T.A., Banks, S., Born, A.F., Bustamante, R.H., Calvopiña, M., Edgar, G.J., Espinoza, E., Fariña, J.M., Garske, L.E., Reck, G.K., Salazar, S., Shepherd, S., Toral-Granda, V., Wallem, P., 2004. A trophic model of a Galápagos subtidal rocky reef for evaluating fisheries and conservation strategies. Ecological Modelling, Placing Fisheries in their Ecosystem Context 172, 383–401. https://doi.org/10.1016/j.ecolmodel.2003.09.019 ↵
- Ortiz, M., Wolff, M., 2002. Spatially explicit trophic modelling of a harvested benthic ecosystem in Tongoy Bay (central northern Chile). Aquatic Conservation: Marine and Freshwater Ecosystems 12, 601–618. https://doi.org/10.1002/aqc.512 ↵
- Piroddi, C., Akoglu, E., Andonegi, E., Bentley, J.W., Celić, I., Coll, M., Dimarchopoulou, D., Friedland, R., de Mutsert, K., Girardin, R., Garcia-Gorriz, E., Grizzetti, B., Hernvann, P.-Y., Heymans, J.J., Müller-Karulis, B., Libralato, S., Lynam, C.P., Macias, D., Miladinova, S., Moullec, F., Palialexis, A., Parn, O., Serpetti, N., Solidoro, C., Steenbeek, J., Stips, A., Tomczak, M.T., Travers-Trolet, M., Tsikliras, A.C., 2021. Effects of Nutrient Management Scenarios on Marine Food Webs: A Pan-European Assessment in Support of the Marine Strategy Framework Directive. Front. Mar. Sci. 8. https://doi.org/10.3389/fmars.2021.596797 ↵
- Piroddi, C., Coll, M., Macias, D., Steenbeek, J., Garcia-Gorriz, E., Mannini, A., Vilas, D., Christensen, V., 2022. Modelling the Mediterranean Sea ecosystem at high spatial resolution to inform the ecosystem-based management in the region. Sci Rep 12, 19680. https://doi.org/10.1038/s41598-022-18017-x ↵
- Romagnoni, G., Mackinson, S., Hong, J., Eikeset, A.M., 2015. The Ecospace model applied to the North Sea: Evaluating spatial predictions with fish biomass and fishing effort data. Ecological Modelling 300, 50–60. https://doi.org/10.1016/j.ecolmodel.2014.12.016 ↵
- Salomon, A.K., Waller, N.P., McIlhagga, C., Yung, R.L., Walters, C., 2002. Modeling the trophic effects of marine protected area zoning policies: A case study. Aquatic Ecology 36, 85–95. https://doi.org/10.1023/A:1013346622536 ↵
- Vilas, D., 2022. Spatiotemporal Ecosystem Dynamics on the West Florida Shelf : Prediction, Validation, and Application to Red Tides and Stock Assessment. University of Florida. ↵
- Walters, C., Christensen, V., Walters, W., Rose, K., 2010. Representation of multistanza life histories in Ecospace models for spatial organization of ecosystem trophic interaction patterns. Bulletin of Marine Science 86, 439–459. ↵
- Christensen, V., Coll, M., Steenbeek, J., Buszowski, J., Chagaris, D., Walters, C.J., 2014. Representing Variable Habitat Quality in a Spatial Food Web Model. Ecosystems 17, 1397–1412. https://doi.org/10.1007/s10021-014-9803-3 ↵
- Steenbeek, J., Coll, M., Gurney, L., Mélin, F., Hoepffner, N., Buszowski, J., Christensen, V., 2013. Bridging the gap between ecosystem modeling tools and geographic information systems: Driving a food web model with external spatial–temporal data. Ecological Modelling 263, 139–151. https://doi.org/10.1016/j.ecolmodel.2013.04.027 ↵
- Coll, M., Steenbeek, J., 2017. Standardized ecological indicators to assess aquatic food webs: The ECOIND software plug-in for Ecopath with Ecosim models. Environmental Modelling & Software 89, 120–130. https://doi.org/10.1016/j.envsoft.2016.12.004 ↵
- Steenbeek, J., Corrales, X., Platts, M., Coll, M., 2018. Ecosampler: A new approach to assessing parameter uncertainty in Ecopath with Ecosim. SoftwareX 7, 198–204. https://doi.org/10.1016/j.softx.2018.06.004 ↵
- Colléter, M., Valls, A., Guitton, J., Gascuel, D., Pauly, D., Christensen, V., 2015. Global overview of the applications of the Ecopath with Ecosim modeling approach using the EcoBase models repository. Ecological Modelling 302, 42–53. https://doi.org/10.1016/j.ecolmodel.2015.01.025[/footnote] [footnote]Coll, M., Akoglu, E., Arreguín-Sánchez, F., Fulton, E.A., Gascuel, D., Heymans, J.J., Libralato, S., Mackinson, S., Palomera, I., Piroddi, C., Shannon, L.J., Steenbeek, J., Villasante, S., Christensen, V., 2015. Modelling dynamic ecosystems: venturing beyond boundaries with the Ecopath approach. Rev Fish Biol Fisheries 25, 413–424. https://doi.org/10.1007/s11160-015-9386-x ↵
- De Mutsert K, Marta Coll, Jeroen Steenbeek, Cameron Ainsworth, Joe Buszowski, David Chagaris, Villy Christensen, Sheila J.J. Heymans, Kristy A. Lewis, Simone Libralato, Greig Oldford, Chiara Piroddi, Giovanni Romagnoni, Natalia Serpetti, Michael Spence, Carl Walters. 2023. Advances in spatial-temporal coastal and marine ecosystem modeling using Ecopath with Ecosim and Ecospace. Treatise on Estuarine and Coastal Science, 2nd Edition. Elsevier. https://doi.org/10.1016/B978-0-323-90798-9.00035-4 ↵
- Walters C, Christensen V, Pauly D. 1997. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Reviews in Fish Biology and Fisheries 7: 139-172. https://doi.org/10.1023/A:1018479526149 ↵
- Christensen V, Walters C. 2004. Ecopath with Ecosim: methods, capabilities and limitations. Ecological Modelling 72: 109-139. https://doi.org/10.1016/j.ecolmodel.2003.09.003 ↵
- Christensen and Walters. 2004. op. cit. ↵
- Walters C, Pauly D, Christensen V. 1999. Ecospace: prediction of mesoscale spatial patterns in trophic relationships of exploited ecosystems, with emphasis on the impacts of marine protected areas. Ecosystems 2: 539-554. https://doi.org/10.1007/s100219900101 ↵
- Walters et al. 1999. op. cit. ↵
- Christensen, V., Coll, M., Steenbeek, J., Buszowski, J., Chagaris, D., Walters, C.J., 2014. Representing Variable Habitat Quality in a Spatial Food Web Model. Ecosystems 17, 1397–1412. https://doi.org/10.1007/s10021-014-9803-3 ↵
- Walters, C., Christensen V, Walters W, Rose K. 2010. Representation of multi-stanza life histories in Ecospace models for spatial organization of ecosystem trophic interaction patterns. Bull. Mar. Sci. 86(2):439-459 ↵
