25 Shared foraging arenas
The basic Ecosim formulation for predation interactions assumes that each non-zero consumption of a prey type i by a predator type j takes place in a foraging arena unique to that interaction[1] [2] [3]. The rationale for this assumption is that each arena is defined by the combined behaviors of both the prey and the predator, and possibly also by selection of particular prey sub-types, (e.g., sizes), such that multiple predators can feed on the same prey type in different ways (at different depths, times of day, spatial microhabitats) without competing directly for prey within the typically very confined space represented by each arena.
The general foraging arena assumption that predation typically is concentrated within restricted arenas, and hence at restricted rates, has profound implications for model predictions about ecosystem stability, and the further assumption that each predator-prey interaction takes place within a unique arena has equally profound implication for the maintenance of ecosystem structure and diversity[4]. It essentially represents the possibility of a distinct "feeding niche" for each of the predators that takes type i prey, hence allowing for the possibility that multiple predators can coexist while feeding on only that prey type. A prototype example of this possibility is with rockfishes (Sebastes spp.) along the Pacific coast, where a diverse collection of species all feed on euphausids, but avoid direct competition for these euphausids by feeding at different depths and times of day. An obvious evolutionary argument in favour of assuming such fine structure in feeding interactions is that if several predators were to feed within the same micro-scale foraging arena, the intense inter-specific competition caused by such behavior would result in very strong natural selection favouring differentiation of behaviors to avoid it, e.g., by feeding at different depths or times.
While there are evolutionary arguments in favour of assuming a distinct foraging arena for every interaction, Aydin and Gaichas[5] emphasize that there are some situations where multiple predator types are likely to feed on exactly the same prey and at the same place and time. An example could be where the predator types represent different life history stanzas (age-size classes) of the same predator species with very similar feeding modes (times and locations).
We represent this possibility in Ecosim by entry of base proportions of each predator type’s diet that occurs in each of the possible foraging arenas defined by all non-zero predator-prey consumption linkages. Vulnerable prey density in each arena is then represented as varying over time in response to abundances of all predator types that feed in the arena.
In Ecosim, we define a list a = 1, … , Na of possible foraging arenas, where Na is the number of non-zero consumption interactions in the Ecopath diet matrix representing consumption of each prey type i by predator type j. Each of these potential arenas has a defining prey type i(a) and defining predator type j(a).
When only predator type j(a) feeds in arena a, vulnerable prey density Va is predicted by the basic foraging arena equation,
[latex]V_a=\frac{v_a \cdot B_{i(a)}}{v_a+v_a' +\alpha_a \cdot P_{j(a)}}\tag{1}[/latex]
Here, va and va' are vulnerability exchange rates of prey to and from arena a, Bi(a) is prey biomass, Pj(a) is predator abundance (biomass or sum of numbers times search rates per predator for multi-stanza predators), and αa is the predator rate of effective search (volume swept per time divided by foraging arena volume). The predation flow rate (biomass of prey i(a) consumed per unit of time by predator j(a)) is then predicted as Qi(a),j(a) = αaVaPj(a). The va and αa are parameterized by having model builders define va from maximum possible mortality rates expressed as multiples Ka of Ecopath base instantaneous predation rates Mij(0) = Qij(0) / Bi(0), simply by setting va = Ka where the superscript (0) designates Q’s and B’s estimated as base (initial) values of abundances and flows in the Ecopath baseline model. The back-exchange parameter v’ is set equal to v since it cannot be estimated separately from the αa parameter.
The shared-arena extension of Eq. 1 is straightforward,
[latex]V_a=\frac{v_a \cdot B_{i(a)}}{v_a+v_a'+ \sum \limits_k \alpha_{ak} \cdot P_k}\tag{2}[/latex]
Here the predator impact on Va is represented by a sum over all possible predators k of arena-specific search rates αak times predator abundances Pk. [In the software-implementation of this, we do not actually sum over all k but instead construct a list of all non-zero αak flow combinations, and sum the αakPk denominator terms only over the elements of that list.]
To parameterize Eq. 2 in a relatively simple way while assuring that it predicts predation rates equal to Ecopath base rates when the system is at its Ecopath base state, we need to specify base proportions pak of each predator k’s diet that is taken in arena a. These proportions are constrained to sum to Ecopath base consumption rates over all a for which i(a) = i. That is, we take the by-arena base flows to be pak Qi(a),k(0). These base flows then imply a base instantaneous mortality rate Ma(0) totaled over predators feeding in a, for prey i(a),
[latex]M_a^{(0)}=\frac{\sum \limits_k Q_{ak}^{(0)}}{B_{i(a)}}\tag{3}[/latex]
Using this input or baseline estimate of M for each arena and an assumed vulnerability multiplier Ka for that arena, we simply set va = Ka (and va' = va).
Next, note that to be consistent with Ecopath baseline inputs, we must require that Ecosim predict Qak(0) when all biomasses (and p’s) are at their Ecopath base values. The Ecosim prediction of rate Qak (flow rate of prey to predator k from feeding in arena a) at any time is Qak = αakVaPk, implying we must constrain the αak so that Qak(0) = αak Va(0) Pk(0), i.e. we must set αak = Qak(0) / (Va(0) Pk(0) ). This means that to estimate the αak we must first estimate the base vulnerable abundances Va(0).
This estimation turns out to be remarkably simple, when we note that the Ecopath base value of ∑k αak Pk must equal ∑k Qak(0) / Va(0), (simply sum Qak over k, which must equal Va = ∑k αak Pk, and solve for ∑k αak Pk). Substituting ∑k Qak(0) / Va(0) for ∑k αak Pk in Eq. 2, then solving for Va(0), we calculate the base vulnerable abundances to be simply,
[latex]V_a^{(0)}= v_a \cdot B_{i(a)} - \frac{\sum \limits_k Q_{ak}^{(0)}}{v_a+v_a'}\tag{4}[/latex]
The αak are then calculated from these base vulnerable biomasses. Time-varying values of Qak are computed efficiently in Ecosim by setting up a list h = 1, … , Nh of all non-zero by-arena flows (Nh ≥ Na), where for each list element we store its associated prey type i(h), predator type k(h), and arena a(h).
To calculate Qak, we sweep down this list repetitively. On the first sweep, we accumulate the denominator sums ∑k αak Pk for Eq. 2. We then sweep down the arena list and calculate Va for every a again using Eq. 2. Then we sweep again down the h list, calculating Qak = αakVaPk and accumulating predictions of total predation rates on the prey i(a) and food consumption rates by predators k(a).
As an added bit of model realism, one can specify a non-zero prey handling times for predator k (type II functional response[6]), and the Qak calculation is modified to be Qak = (αak/Hk) Va Pk, where Hk is the denominator of Holling's multi-species disc equation for predator k feeding. This handling time correction is also applied in the bout-feeding formulation described in the next chapter.
To edit the pak diet proportions array, we display a matrix for each prey type i of the non-zero i-k consumption proportions, as shown schematically in Table 1. In this table, m is the number of non-zero flows from prey ito predators k where each such flow defines a potential foraging arena. Note that each column of the table must sum to 1.0, i.e. all of the consumption by predator kj of prey type i must be accounted for by feeding in one of the m identifiable arenas for prey type i. The Ecosim default proportions for this table imply that each predator takes all of its consumption of prey type i in a unique arena, i.e. the table is an identity matrix, (with values of 1 on the shaded diagonal in Table 1).
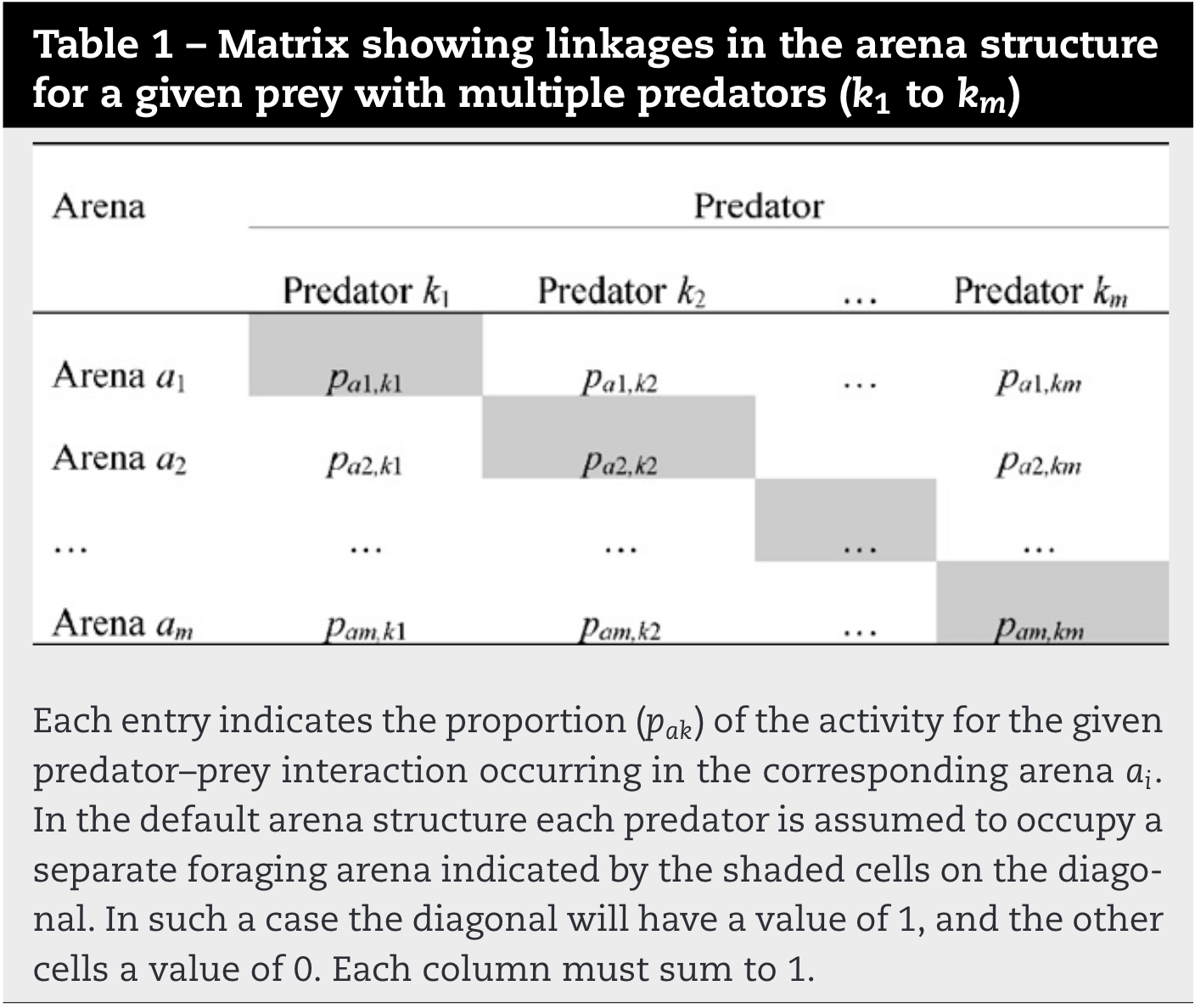
The opposite extreme of this default assumption would be that all consumption of prey type i by its predators occur in only one arena or behavioral state for prey i, as shown in Table 2. This case implies maximum possible impact of predators k on availability of prey i to one another, and will cause competitive exclusion of at least some predator types in Ecosim unless the predators are well-differentiated in terms of overall diet composition, i.e. where each predator "specializes" on a different prey type i, which dominates the diet composition, as for instance shown by Schmidt[7]. Studies rather tend to indicate resource partitioning between competing predator species, leading to non-additive mortality rates, see e.g., Griffen and Byers[8]. Separation where diet compositions indicate predator overlap may also be caused by temporal exclusion of prey based on availability to the predator[9].

In the special case where a set of predators feeds on only one prey type in a single arena (Table 2), and where there are no complications such as multistanza population dynamics where abundance of one or more predator types may be limited by recruitment rates from younger stanzas, the above formulation implies that there is not even a unique equilibrium point for predator abundances. Rather, all predator abundance combinations that predict V=V(0) in Eq. 2 are neutral stable points provided predator mortality rates remain at Ecopath base values, such that any temporary pulse of differential mortality that causes one or more predators to decline will then be followed by persistence of the new predator abundance combination if mortality rates return to the base values. Any predator that suffers a persistent differential increase in mortality rate is predicted to decline toward extinction.
Attribution This chapter was inspired by Kerim Aydin’s work on a foraging arenas, and is based on Walters and Christensen. 2007.[10], used with permission from Elsevier, Licence Number 5663310244809.
- Walters, C., V. Christensen and D. Pauly. 1997. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Reviews in Fish Biology and Fisheries 7:139-172. https://doi.org/10.1023/A:1018479526149 ↵
- Walters, C.J., J.F. Kitchell, V. Christensen and D. Pauly. 2000. Representing density dependent consequences of life history strategies in aquatic ecosystems: Ecosim II. Ecosystems 3: 70-83. https://doi.org/10.1007/s100210000011 ↵
- Christensen, V. and C. J. Walters. 2004. Ecopath with Ecosim: methods, capabilities and limitations. Ecol. Model. 172:109-139 https://doi.org/10.1016/j.ecolmodel.2003.09.003 ↵
- Walters, C. J. and Martell, S. J. D., 2004. Fisheries Ecology and Management. Princeton University Press, Princeton. 399 pp. ↵
- Aydin, K. Y. and Gaichas, S. K., 2007. In defense of complexity: towards a representation of uncertainty in multispecies models. MS, SC/58/E, Alaska Fisheries Science Centre, NOAA, Seattle WA ↵
- Holling, C.S., 1959. The components of predation as revealed by a study of small mammal predation of the European pine sawfly 91, 293–320. https://doi.org/10.4039/Ent91293-5 ↵
- Schmidt, K. A., 2004. Incidental predation, enemy-free space and the coexistence of incidental prey. Oikos, 106:335-343. https://doi.org/10.1111/j.0030-1299.2004.13093.x ↵
- Griffen, B. D. and Byers, J. E., 2006. Partitioning mechanisms of predator interference in different habitats. Oecologia, 146:608-614. https://doi.org/10.1007/s00442-005-0211-4 ↵
- Scheuerell, J. M., Schindler, D. E., Scheuerell, M. D., Fresh, K. L., Sibley, T. H., Litt, A. H. and Shepherd, J. H., 2005. Temporal dynamics in foraging behavior of a pelagic predator. Canadian Journal of Fisheries and Aquatic Sciences, 62:2494-2501. https://doi.org/10.1139/f05-164 ↵
- Walters, C and V. Christensen. 2007. Adding realism to foraging arena predictions of trophic flow rates in Ecosim ecosystem models: shared foraging arenas and bout feeding. Ecological Modelling 209:342-350. https://doi.org/10.1016/j.ecolmodel.2007.06.025 ↵
