Module 4: Tropical Forest Plant Growth Forms as Related to Natural Forest Management
Topic 4.1: Trees
Because of their large sizes and great longevities, trees have tremendous effects on the ecology of tropical forests. Tropical trees are also important at the planetary level; carbon stored in their trunks and roots is important in maintaining global carbon balance and in slowing global warming and sea level rise. At the level of large watersheds like the Amazon, it is important to recognize that a large portion of the rain falling on forests is derived from water transpired by trees. At a more local level, light rains may never reach the forest floor, the water instead being intercepted by and evaporated from tree leaves and branches. Trees also provide structural support for a variety of other growth forms, such as epiphytes and vines. Although knowledge about other types of plants is important when trying to manage forests sustainably, we obviously need to know a lot about how trees function, starting with below-ground processes.
The root systems of tropical trees vary a great deal by species and with environmental conditions (e.g., depth to mechanically impeding horizons, bedrock, or the water table). Tropical forests are famous for their diversities of aerial roots, buttressed roots, apogeotropic (=upward growing) roots, and pneumatophores (=so-called “breathing” roots). But most species of tropical trees have roots that are similar to those of trees in temperate forests in both structure and distribution.
Although most fine roots of tropical trees are generally concentrated near the soil surface where nutrients are more available, deep roots also develop where conditions allow. These deep roots may be particularly important for water uptake when surface soils have dried; the discovery of roots at depths of 10-14 m in Amazonia suggests that roots may be exploiting soil resources substantially deeper than typical samples taken by soil scientists.
The horizontal spread of tree roots has not been much studied in the tropics but, like rooting depth, most likely varies by species and site conditions. That roots extend well beyond the crown margins of many trees is abundantly clear. Even at the seedling stage, horizontal root growth can be substantial. Allowing the roots of tree seedlings to expand freely is often problematic in nurseries where “pot-bound” plants are all too common. Out-planted seedlings can also suffer the same “pot-bound” problem when set out in compacted soils.
Roots proliferate in resource-rich volumes of soil. Given that the distributions of nutrients and water in soils are extremely heterogenous, fine root densities also can vary substantially over even very short distances. And these distributions change over time, sometimes very rapidly as when an animal dies or defecates, creating thereby a local, if temporary, concentration of nutrients.
Maintenance of the appropriate balance between root surface area for absorption and leaf surface area for photosynthesis and transpiration is critical for tree survival and growth. As environmental conditions change seasonally or due to silvicultural practices, so must this so-called “root-shoot ratio.” For example, because evapo-transpirational demands in the understory are low, understory saplings tend to have relatively low root-shoot ratios. When the canopy is opened due to logging or thinning, these saplings must rapidly adjust their root-shoot ratios for the new conditions of higher light intensities, higher temperatures, and higher vapor pressure deficits. For the same reason, nursery-grown seedlings are often intentionally “hardened up” by gradual deprivation of water and nutrients before out-planting so as to reduce “transplant stock.” Pruning off a portion of leaf area of these seedlings can also help to reduce transplant shock by increasing root-shoot ratios. Pre-planting shoot pruning is made even more critical when seedlings are purposefully or inadvertently root-pruned to facilitate lifting and planting.
Little is known about the long-term impacts on tropical trees of root damage, but enough is known about temperate oaks (Quercus spp.) and other hardwoods to raise concern. Root damage is common where ground-based timber yarding operations are carried out, particularly if they continue during wet weather. But which species are most susceptible to root-invading pathogens? Do trees that suffer mechanical root damage subsequently develop butt and heart rots? Do trees with damaged roots suffer elevated mortality rates and decreased growth rates? Research is needed to answer these and other questions, but in the meantime, logging damage to roots needs to be minimized.
Roots of virtually all species of tropical trees are symbiotically associated with fungi that assist in nutrient uptake. Very fine hyphae (=fungal threads) of the fungi attached to tree roots grow out into the soil, thereby greatly increasing the host trees’ below-ground surface area and rates of uptake of nutrients, especially of phosphorous. The fungi benefit from the relationship primarily by receiving sugar from their host tree. There are different sorts of these mycorrhizal associations (e.g., endo- and ectomycorrhizae) involving many different species of fungi. Generally the spores and other inocula of these fungi are abundantly available in tropical forest soils. Only where species are newly introduced outside of their natural geographical range, on extremely degraded soils, and in nurseries is lack of mycorrhizae sometimes an issue. Generally by incorporating forest soil into planting mixes, this problem can be avoided.
Due to their commercial importance for timber and the fact that they grow above ground where we can see them, tree stems have received more attention from researchers than tree roots. Among tropical trees are species with typically fluted, anastomosing, locally swollen, and other variations on the basic tapered cylinder. Tree stems are subjected to abiotic stresses (e.g., wind damage and fire) and biological attacks (e.g., entwining vines and heartrot fungi) that affect their marketability. Stem quality, from a timber production perspective, is generally recorded in permanent plots and during inventories using a classification system such as that presented in Figure 3-1 (stem form classes, crown classes). Note, for example, that if a primary objective of forest management is maintaining an increasing populations of animals that use tree cavities for nesting or denning, these “quality” classes might be arranged quite differently.
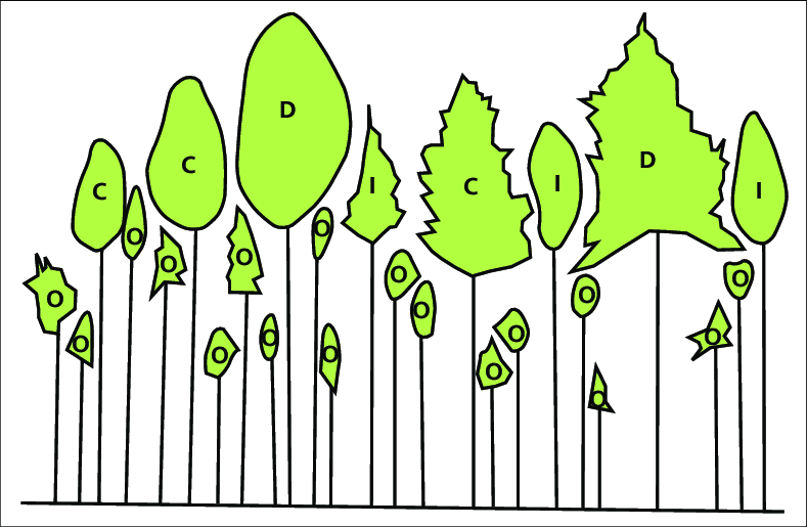
Figure 4.1.1. A common crown class system in which D = dominant, C = co-dominant, I = indirect light (or subdominant), and O = overtopped. Sometimes a fifth class is included for trees that receive substantial illumination from the side.
Characteristics of tree crowns such as width and depth serve as good indicators of the local conditions under which a tree has been growing and may also be useful in predicting a tree’s response to thinning. Broad and deep crowns indicate, for example, that the tree had plenty of growing space. Trees with small or incomplete crowns (Figure 3-1) typically do not respond well to release treatments due to having been suppressed for too long. When the canopy surrounding such a tree is radically opened, it may die or at least take a very long time to respond. This poor reaction is likely due to some combination of having a root-shoot ratio inappropriate to the new conditions, mechanical instability when neighbors are removed, and bark scald.
A more detailed examination of a tree’s branching patterns can reveal even more about its history and even its wood properties. The duration of branch retention and the process by which they are finally shed, for example, influence the knottiness of wood and whether the knots are loose or tight. The high value of knot-free or “clear” timber can sometimes justify branch pruning in intensively managed stands. Note that when pruning it is always preferable to remove branches while they are small. A rule of thumb is that branches >30% of the diameter of the main trunk themselves function as trunks, i.e., they have lost the characteristic branch collar at the base of smaller branches that helps keep decay from penetrating into the main stem from the branch stub.
Bark is the first line of defense of a tree stem against mechanical damage and attack by herbivores and pathogens. The often high concentration of extractives (e.g., resins and gums) as well as the abundance of long “bast” fibers in bark constitute part of this defense of the vascular cambium and xylem. Bark also protects the cambium from high temperatures during fires; predictions of tree survival through controlled burns, for example, are often based on bark thickness. Direct solar radiation on the stems of previously shaded can also cause damage if the bark is not thick or rough enough to protect the cambium from the heat load. Generally 60C is taken to be the lethal temperature in the cambium.
Vascular cambia vary seasonally in their activity cycles and hence in their susceptibility to damage. When the cambium is active, for example, bark is more easily peeled by herbivores and also inadvertently by timber yarding machinery. Some tree species produce successive cambia and hence develop included phloem (layers of phloem alternating with layers of xylem). Whether this form of developmental anatomy in trees confers resistance to or capacity to recuperate from damage has apparently not been determined in trees but seems important in some woody vines (see below). Due to the substantial potential long-term negative consequences of even minor mechanical damage, protecting trees during forest management operations is obviously critical.
Trees respond to damage by depositing defensive compounds around the damaged tissues, thereby compartmentalizing the decay. Trees therefore do not “heal” in the same sense as animals, but they can restrict the proliferation of decay. Living wood cells (parenchyma) must be present near the site of damage for compartmentalization to occur. Due to scarcity of living parenchyma in the heartwoods of many tree species, the compartmentalization potential is limited and the likelihood of heartrots is increased. A few decades after destructive logging operations or ground fires, 50% or more of the large trees may be hollow or have heartrots.
Many silvicultural operations are applied to optimize the leaf areas and crown exposures of potential crop tree. It is important to recognize that all leaves are not constructed the same and that during the life span of a single leaf (ranging from several months to many years), physiological characteristics change as well. Leaves that develop where light intensities are high, for example, are typically thick, small, covered with a thick cuticle, and held at a steep angle to horizontal. Shade leaves, in contrast, tend to be larger, thinner, and darker green due to a higher concentration of chlorophyll. Based on these differences between “sun” and “shade” leaves, it is easy to understand why some trees must shed and replace their leaves before they can respond favorably to canopy-opening treatments. As leaves age and become encrusted with lichens, algae, and other epiphylls, their photosynthetic capacity declines.