Module 5: Plant and Forest Ecosystem Water, Carbon, and Nutrient Relations
Topic 5.3: Nutrient Relations
A great deal has been written about soil nutrients and how they are taken up by plants; scores of studies on this topic are published every month. Rather than try to summarize this extensive field, this section will focus on some of the pitfalls and problems confronting tropical foresters who get involved in soil and plant nutrient analysis. Sophisticated techniques for soil and leaf tissue analyses are now available, but the data they provide are often difficult to interpret. The basic problems with studying plant nutrient relations are that nutrient availability is spatially extremely variable and the quantities of nutrients actually available to plants may differ substantially from the nutrients measured in a soil analysis laboratory.
Before elaborating on some of the challenges associated with assessing availability of nutrients to plants, perhaps it would be useful to review which nutrients are needed and how plants obtain them. We will concentrate on the nutrients that most often limit plant growth, but remember that under some circumstances, micronutrient deficiencies and other imbalances can be challenging but there are some typical responses (Figure 5.3.1).
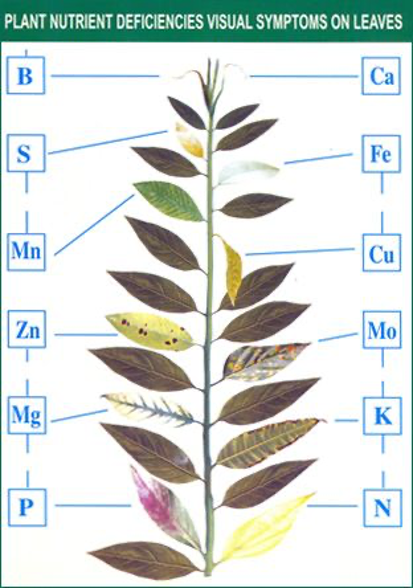
Figure 5.3.1. Indicators of nutrient and micronutrient deficiencies
Of the 27 elements required for plant growth, two are most often limiting for terrestrial plants: nitrogen and phosphorus. Providing even the briefest overview of these nutrients is challenging insofar as they are so different. For example, nitrogen is the most abundant gas in our atmosphere but needs to be converted (=”fixed”) into a form available to plants by microbes, lightning, or industrial processes. Due to the activities of nitrogen-fixing organisms, some of which are associated with plant roots (e.g. Rhizobium bacteria in some Leguminosae, Frankia actinomycetes in some Rhamnaceae, Betulaceae, and Casuarinaceae), nitrogen is less often in short supply in mature soils than on soils recently derived from parent rocks, wind-blown sand, volcanic ash, or alluvial material. Phosphorus, in contrast, is derived mostly from weathered rocks and most often is limiting on old, heavily leached soils. Another difference is that nitrogen compounds (e.g., NO3– and NH4+) are fairly soluble in soil water, whereas phosphorus tends to be found in forms unavailable to plant roots. Due to the low solubility of phosphorus compounds, most plants depend on uptake by the fine hyphae of symbiotic fungi associated with their roots (mycorrhizae) to obtain sufficient phosphorus.
Assessing soil nutrient availability to trees and other long-lived and large plants is made complicated by the volumes of soil they exploit, the spans of time over which they extract nutrients, their great storage capacities, and the various effects that the plants themselves have on nutrient distribution and availability. To begin to understand this problem it first helps to disregard several commonly held ideas about tropical trees: that they are shallowly rooted and that their lateral roots extend about as far as their branches. Lateral roots often grow far beyond the crown edge, and, like trees anywhere, tropical trees are deeply rooted when soil conditions allow but are shallowly rooted where rocks or anaerobic conditions due to soil flooding preclude deep rooting. Although fine roots are concentrated near the surface where nutrients are released from leaf litter, roots of some trees, woody vines, and palms may penetrate 10 m or more where soil conditions allow. Estimating soil nutrient availability for a plant with 10 m deep roots by sampling surface soils is obviously fraught with problems.
Soil nutrients are also not evenly distributed in space or time. Although nutrient concentrations generally decrease with depth, at any one depth they may vary substantially over distances of less than one meter. For example, pockets of organic matter (e.g., dung piles or leaf litter stored in animal burrows), might hold and release in plant-available forms five times more nitrogen, calcium, and other nutrients than a few centimeters away in the soil. Plant roots respond to this heterogeneity by proliferating in nutrient-rich soil volumes, and growing rapidly through volumes where nutrients are limiting. To help account for spatial heterogeneity, individual soil samples are often combined (=bulked) before analysis to obtain a composite picture of soil nutrient concentrations.
Understanding what happens at the interface between a fine root or mycorrhizal hyphae and the soil is critical for understanding what proportions of total soil phosphorus, nitrogen, and other nutrients are available for plant uptake. Roots change the soil around them by exuding mucus and other sugar-based compounds that lubricate root passage through the soil and provide food for bacteria and other microbes that proliferate in the rhizosphere. Microbial concentrations, measured in millions of organisms per cm3 of soil, can be 100 times higher within 1 mm of the tip of a growing root than a few centimeters away in the soil. These microbes can greatly modify nutrient availability to plant roots, and some are non-symbiotic nitrogen fixers. Plant roots also exude compounds that change soil pH or attach to nutrient ions to form compounds that are more readily absorbed. It is difficult to mimic this “chelation” effect in a soil laboratory, but the pH of soil samples can be manipulated so as to mimic the conditions at the root-soil interface.
To measure available nutrients in a soil sample, the nutrients first need to be extracted into an aqueous solution. But what are the appropriate solutions and other conditions under which the nutrient extractions should be conducted? Soil pH, in particular, can greatly affect the process of nutrient ion adsorption and release from soil particles. Organic matter, often the principal reservoir of soil nutrients, is especially sensitive to the pH of the extraction solution. For example, estimated concentrations of available phosphorus (ug/g) in a soil sample extracted with a neutral solution would be different than extraction with a buffered solution at pH9 or with a weak acid at pH5. But what is the appropriate pH?
To get around this problem of determining what type of extraction solution is appropriate, soil scientists often use what are called “ion exchange” methods. The basis for these methods is that plant roots and mycorrhizal hyphae only take up nutrients that are in the soil solution. Nutrient ions can be weathered from rocks, mineralized from decomposing organic matter, or released from cation or anion exchange sites on small particles of organic matter or clay micelles. Sheets of ion exchange resins to which some ions readily attach that are inserted into the soil take up nutrients as they enter the soil solution. In the laboratory the quantities of absorbed ions can be determined.
Alternatives to chemical extraction and ion exchange methods for assessing soil nutrient availability is to conduct bioassays using fast-growing plants, or to conduct fertilizer addition experiments. For bioassays, most researchers plant vegetable seeds (e.g., spinach) in pots filled with the soil to be assessed; nutrient limitations can then be determined on the basis of nutrient deficiency syndromes (Figure 5.3.1), or by comparing growth rates in soils to which nutrients have been augmented element by element. Nutrient addition experiments can also be conducted in the field, but in either case, the form in which the nutrients are added (e.g., NO3– or NH4+ supplying compounds) can affect the results. Researchers should be warned that due to their great capacity for “luxury consumption” and storage, assessing nutrient limitations in trees with fertilizer experiments may require several years to yield definitive results.
For well-known species, foliar nutrient analysis can be used to reveal deficiencies. When sampling leaves it is important to control for leaf age and crown position because both can influence nutrient concentrations.