Module 5: Plant and Forest Ecosystem Water, Carbon, and Nutrient Relations
Topic 5.1: Water Relations
Water plays many roles in plants: it is the primary fluid in which biochemical reactions occur; water under positive pressure in cells (turgor) provides mechanical support for leaves and other herbaceous plant parts; and, water is used for transpirational cooling. Actually, 90% or more of the water that moves through the soil-plant-atmosphere continuum (SPAC) is used for cooling. And the quantities of water used by large trees are prodigious; a 50 cm dbh tree in moist soil, for example, may transpire 100-200 l of water in a single day. Given the heat load of sunshine, the narrow temperature range over which photosynthetic and other enzymes function, and the consequences of over-heating (membrane damage, protein denaturation, and death), the importance of water for evaporative cooling is abundantly clear. Along the SPAC through the roots and stem, and out through the leaves, there are numerous variable and fixed resistances to water flow. As water enters the roots it encounters resistances due to having to move through intercellular spaces in the cortex and through living cells in the endodermis. Most of the flow occurs in hollow tubes in the xylem (vessels and tracheids) that cause resistances that decrease with increasing aperture diameter (flow increases with vessel radius to the fourth power!) and vessel length. At the other end of the plant, up in the leaves, the opening and closing of the stomatal guard cells provides the largest variable resistance to water flow. Water vapor that exits a leaf through a stomatal aperture encounters the “boundary layer” resistance, the zone of still air around the surface through which water (and carbon dioxide) moves by the slow process of molecular diffusion rather than by the more rapid process of convection (i.e., air turbulence). The larger the leaf and the farther from the leaf margin, the thicker the boundary layer. It is worth considering in detail how water moves through plants because lack of water is the most common cause of slow growth and mortality, especially among recently established or transplanted seedlings. Water deficits occur primarily because transpiration occurs more rapidly than water can be replenished in plant tissues. Dry soil is the most obvious cause of this condition but plants can suffer water deficits in moist soil if their root-shoot ratio is too low or their transpiration rates are excessive. Mid-day leaf wilt, for example, can indicate that due to resistances along the SPAC, water necessary for maintaining leaf turgor was temporarily insufficient. But it should also be noted that by reducing the leaf area held perpendicular to the rays of the sun, temporary wilting may help leaves avoid high heat loads. Soil compaction, by reducing root growth rates and therefore the volume of soil exploited by a plant’s root system, can also result in plant water deficits even under moist condition.
The primary and most rapid response to excessive water loss is stomatal closure. With the stomates closed, water loss still occurs through the epidermis, but at a much slower rate. Stomatal closure also halts uptake of carbon dioxide and therefore stops photosynthesis (except in some cacti, orchids, and other succulents that store carbon dioxide at night for use during the day). Leaves with closed stomates are also likely to heat up, but the likelihood of tissue damage due to water loss must make stomatal closure worth the temperature risk.
Prolonged or seasonal exposure to water deficits often causes leaf shedding and development of leaves that are physiologically and morphologically better adapted to drought. Dropping leaves may be “costly” to the plant in terms of reduced photosynthetic capacity and lost nutrients that will later have to be replaced, but by decreasing the total surface area, water balance can be maintained. The leaves of plants growing in dry environments, for example, tend to be smaller and covered with a thicker cuticle than well-watered plants. Root-shoot ratios of droughted plants also increase (if they survive the stress), which helps them to avoid future water deficits especially if rooting depth also increase.
Flooding and anaerobic conditions
At the opposite extreme from water deficits, flooding damages non-adapted plants due to oxygen deprivation in the roots. Actually the sequence of changes in the soil after flooding are complex, including development of potentially toxic concentrations of aluminum, reduction of sulphur to hydrogen sulfide, and production of methane. Anaerobic conditions (lack of oxygen) in the soil also affects nutrient availability (by reducing decomposition and mineralization rates) and nutrient uptake (by restricting root growth.) To avoid fermentation in root tissues and the damaging effects of anaerobic conditions, plants adapted to flooding increase oxygen availability in roots and often in the rhizosphere (the soil immediately surrounding the roots) by facilitating its downward movement by producing air filled pathways in the root through which oxygen diffuses relatively rapidly. Structures such as multiple stems, fluted or buttressed bases, prop roots, and various sorts of pneumatophores (i.e., breathing roots; upward-growing extensions from lateral roots) all increase the surface area near the flood line into which oxygen can diffuse. On water-logged soils, long-term anaerobic conditions generally results in shallow-rooted trees that are susceptible to uprooting.
Measuring plant water stress and water availability in soils
Although leaf wilting and abscission are obvious indicators of water stress, there are more sophisticated ways of measuring plant water status (Figure 5.1.1 pressure bomb and porometer). Rates of water loss through stomates can be estimated with a diffusion porometer, and xylem water potential can be estimated with a pressure chamber. The latter instrument is elegant in its simplicity, and gives an indication of the tension at which water is held in plant tissues; high tensions (i.e., low water potentials) indicate water stress. Monitoring midday and pre-dawn water potentials can provide indications of water deficits less extreme than those that cause leaves to wilt or be shed.

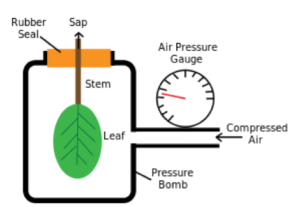
Figure 5.1.1. A pressure chamber used to measure the tension with which water is held in leaves as in indicator of water stress.
Soil water availability can also be measured, but interpretation of the data is generally complicated. Simply by extracting a soil sample and measuring its fresh and oven-dry weights (80o C), percent moisture content (% of dry weight) can be measured. In comparisons of the same soil at different times or different sites with similar soils, percent moisture content data are very useful. Determining how much of the soil water is actually available to plants is more difficult because it varies with both soils and plant species. For example, deep-rooted plants have access to water sources not available to plants that are shallow rooted. A more complicated source of variation in soil water availability at the same percent moisture content is related to soil texture. For example, fine-textured soils (e.g., clays) hold a great deal of water at high tensions which is not available to plants. Soil moisture measurements based on electrical conductivity of the soil solution present the same problem in interpretation. With some more technically sophisticated methods, such as time-domain reflectrometry, some of these problems can be circumvented, but the device still has to be calibrated against moisture release curves from the soil being measured.