Maintenance of the Body
Unit 3: The Lymphatic System, Resistance & Immunity
Unit outline
Part 1: Blood Cell Lineages: Leukocytes
- Characteristics of Leukocytes
- Classification of Leukocytes
Part 2: The Lymphatic and Immune Systems
- Anatomy
- Primary Lymphoid Organs and Lymphocyte Development
- The Organization of Immune Function
- Barrier Defenses and the Innate Immune Response
- The Adaptive Immune Response
- The Cellular Basis of Immunological Memory
- Active versus Passive Immunity
- Diseases Associated with Depressed or Overactive Immune Responses
Part 3: Blood Typing
- Antigens, Antibodies, and Transfusion Reactions
- The ABO and Rh Blood Groups
- Determining Blood Types
Learning Objectives
At the end of this unit, you should be able to:
I. Specify the types of leukocytes, their functions, origins and relative quantities in normal blood.
II. Describe the purpose of a differential count and how to interpret the results.
III. Describe the major functions and anatomical organization of the lymphatic system.
IV. Distinguish between the cardiovascular system and the lymphatic system.
V. Explain nonspecific (innate) resistance to disease and specify the general components of nonspecific (innate) resistance.
VI. Explain specific (adaptive) resistance to disease (immunity), and distinguish between T-cell mediated (cellular) immunity and B-cell mediated (humoral) immunity.
VII. Specify the ways in which antibodies destroy or inactivate a foreign substance in the body.
VIII. Discuss the relationship between antibodies and immunization and specify four ways of conferring immunity.
IX. Describe conditions that may result due to a compromised immune system.
X. Describe the basis of the ABO blood groups and Rh factor and explain the significance of this to transfusions and hemolytic disease of the newborn.
Learning Objectives and Guiding Questions
At the end of this unit, you should be able to complete all the following tasks, including answering the guiding questions associated with each task.
I. Specify the types of leukocytes, their functions, origins and relative quantities in normal blood.
- List all the types of leukocytes, along with their relative frequencies (most common, least common, etc.).
- List and describe in detail the function(s) of the five types of leukocytes. Be as specific and detailed as possible, i.e. include every subtype of leukocyte in your answer!
II. Describe the purpose of a differential count and how to interpret the results.
- What is a differential count? What information can it provide?
- Why are neutrophils found in high numbers in people recovering from burn injuries?
- What could be consequences of an abnormally low neutrophil count?
- How does one of the functions of eosinophils explain their high counts in individuals fighting a parasitic worm infestation?
- What would the basophil count be in people experiencing allergies? Explain why.
- Why is a low lymphocyte count observed in individuals with an active HIV infection? 9. Why is this a dangerous situation?
- Bone marrow disorders cause a low monocyte count. Why?
III. Describe the major functions and anatomical organization of the lymphatic system.
- For each of the following components of the lymphatic system, state its major function(s) and describe its location(s) in the human body:
- Lymphatic vessels
- Lymph
- Primary lymphoid organs
- Secondary lymphoid organs
- Describe the anatomical relationship (i.e. where they are located relative to each other) between lymphatic vessels, lymph, and lymph nodes.
IV. Distinguish between the cardiovascular system and the lymphatic system.
- Compare and contrast the lymphatic and cardiovascular systems by describing:
- Any function(s) that both systems serve.
- Differences between the functions of the two systems.
- Similarities in the overall structure (cell types, tissue types, organs) of the two systems.
- Differences between the overall structures found in the two systems.
V. Explain nonspecific (innate) resistance to disease and specify the general components of nonspecific (innate) resistance.
- What are the main mechanisms that provide the human body with nonspecific resistance to infection?
- Describe the physical and chemical mechanisms used by each of the following tissues to provide the body with a barrier to disease:
- The skin
- Mucous membranes
- Compare and contrast the physical mechanisms used by the skin to provide the body with a barrier to disease with those used by mucous membranes.
- Compare and contrast the chemical mechanisms used by the skin to provide the body with a barrier to disease with those used by mucous membranes.
- Describe the mechanisms by which each of the following acts to provide innate defenses against disease:
- Phagocytes
- Natural killer cells
- Inflammation
- Soluble mediators
- Fever
VI. Explain specific (adaptive) resistance to disease (immunity), and distinguish between T-cell mediated (cellular) immunity and B-cell mediated (humoral) immunity.
- Compare and contrast B-cell mediated (humoral) immunity and T-cell mediated (cellular) immunity, including:
- The stem cells and progenitor cells required
- The location(s) where the cells mature
- The mature cell types involved
- The mechanism by which each mature cell type responds to an antigen
- Describe the major immune functions of phagocytes.
- Describe the immune functions of antigen-presenting cells (APCs).
- List the specific human cell types that can perform phagocytosis, and those that can become antigen-presenting cells. Explain any overlap between your two lists.
- Describe the interactions that occur between the cells of the humoral immunity pathway and cells of the cell-mediated immunity pathway.
- Describe how the human body normally produces antibodies upon exposure to an antigen.
- Compare and contrast the innate and adaptive immune responses in terms of timing, specificity, structures and cells involved, and mechanisms of action. Include advantages and disadvantages of each.
VII. Specify the ways in which antibodies destroy or inactivate a foreign substance in the body.
- Explain each of the 4 possible ways in which an antibody can interact with an antigen to reduce or prevent damage caused by a pathogen (including the mechanism(s) through which each way functions to destroy or inactivate a pathogen or toxin).
VIII. Discuss the relationship between antibodies and immunization and specify four ways of conferring immunity.
- Discuss the premise of conferring immunity through vaccination.
- Briefly describe each of the following methods of conferring immunity:
- Live attenuated vaccines
- Killed inactivated vaccines
- Toxoid vaccines
- Antibody therapy
IX. Describe conditions that may result due to a compromised immune system.
- Describe the immunological basis of immunodeficiencies in general, and differentiate between inherited and acquired immunodeficiencies. Briefly describe one example of each (i.e., identify the deficiency for the inherited example, and the cause and eventual deficiency of the acquired example).
- Describe the immunological basis of autoimmune diseases in general, and name and briefly describe at least two specific examples of autoimmune diseases.
X. Describe the basis of the ABO blood groups and Rh factor and explain the significance of this to transfusions and hemolytic disease of the newborn.
- List all the antigens and antibodies present shortly after birth in each of the 8 major blood types found in humans.
- Create a chart with all possible human blood types that shows which blood types may donate to which others, and which may not. Explain any cases where one type may receive erythrocytes from, but not donate to, another specific blood type (e.g. “blood type X may not donate to blood type Z, but blood type Z could donate to blood type X”).
- Explain the conditions under which anti-D antibodies are produced in humans.
- Describe how hemolytic disease of the newborn occurs and how it can be prevented.
Part 1: Blood Cell Lineages
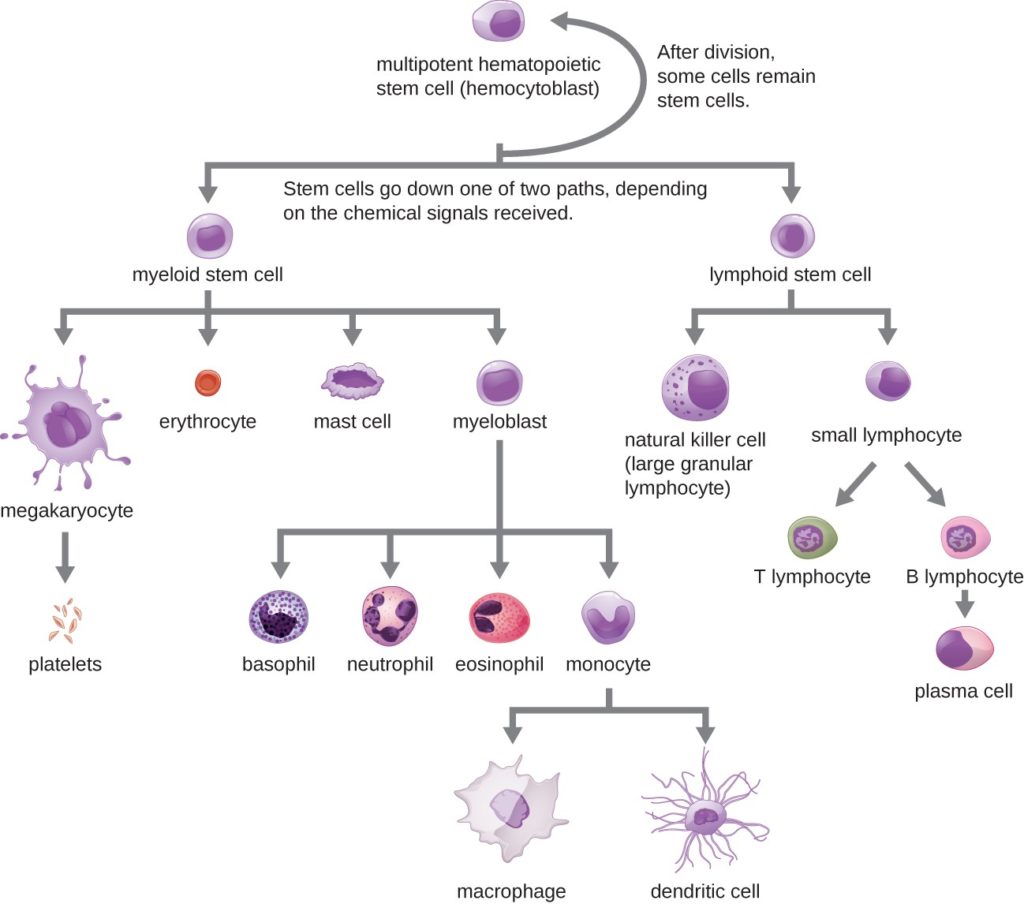
The cells of the blood, including all those involved in the immune response, arise in the bone marrow via various differentiation pathways from hematopoietic stem cells (Figure 1). In contrast with embryonic stem cells, hematopoietic stem cells are present throughout adulthood and allow for the continuous differentiation of blood cells to replace those lost to age or function.
Leukocytes: The leukocyte, commonly known as a white blood cell (or WBC), is a major component of the body’s defenses against disease. Leukocytes protect the body against invading microorganisms as well as genetically transformed body cells that are potentially cancerous. They also clean up extracellular debris and can signal and enhance the healing and repair process.
Characteristics of Leukocytes: Although leukocytes and erythrocytes both originate from hematopoietic stem cells in the bone marrow, they are very different from each other in many significant ways. For instance, leukocytes are far less numerous than erythrocytes. Typically, there are only 5000 to 10,000 leukocytes per microliter (µl) of blood compared to the roughly 5 million erythrocytes. They are also larger than erythrocytes, possessing a nucleus and organelles while erythrocytes expel these structures early in development. Although there is just one type of erythrocyte, there are many types of leukocytes. Most of these leukocytes have a much shorter lifespan than that of erythrocytes, some as short as a few hours or even a few minutes in the case of acute infection.
One of the most distinctive characteristics of leukocytes is their movement. Whereas erythrocytes spend their days circulating within the blood vessels, leukocytes routinely leave the bloodstream to perform their defensive functions in the body’s tissues. For leukocytes, the vascular network is simply a highway they travel and soon exit to reach their true destination. When they arrive, they are often given distinct names, such as macrophage or microglia, depending on their function.
Once they have exited the capillaries, some leukocytes will take up fixed positions in lymphatic tissue, bone marrow, the spleen, the thymus, or other organs. Others will move about through the tissue spaces (diapedesis), very much like amoebas, continuously extending their plasma membranes, sometimes wandering freely, and sometimes moving toward the direction in which they are drawn by chemical signals. This attracting of leukocytes occurs because of positive chemotaxis (literally “movement in response to chemicals”), a phenomenon in which injured or infected cells and nearby leukocytes emit the equivalent of a chemical “911” call, attracting more leukocytes to the site. In medicine, determining the quantity of the different leukocytes can provide pertinent clinical information. These differential counts of the types and percentages of leukocytes present in a sample are often key indicators in making a diagnosis and selecting a treatment.
Classification of Leukocytes: When scientists first began to observe stained blood slides, it quickly became evident that leukocytes could be divided into two groups, according to whether their cytoplasm contained highly visible granules:
- Granular leukocytes contain abundant granules within the cytoplasm. They include neutrophils, eosinophils, and basophils.
- While granules are not totally lacking in agranular leukocytes, they are far fewer and less obvious. Agranular leukocytes include monocytes, which mature into phagocytic macrophages, and lymphocytes, which arise from the lymphoid stem cell line.
Granular Leukocytes: We will consider the granular leukocytes in order from most common to least common. All of these are produced in the red bone marrow and have a short lifespan of hours to days. They typically have a lobed nucleus and are classified according to which type of stain best highlights their granules (Figure 2).
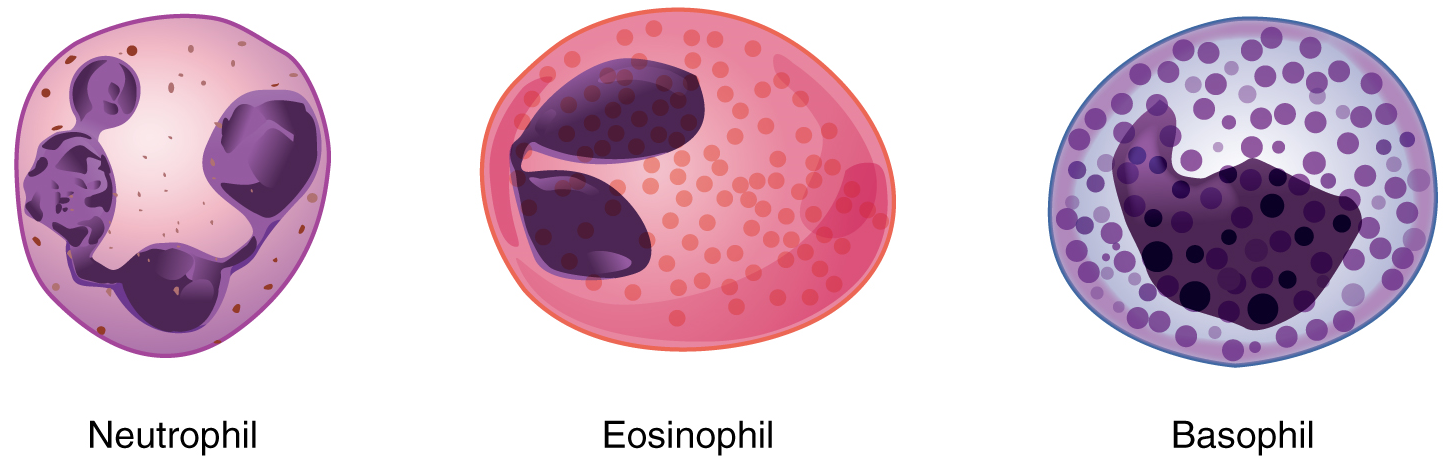
The most common of all the leukocytes, neutrophils will normally comprise 50–70 percent of total leukocyte count. They are called neutrophils because their granules show up most clearly with stains that are chemically neutral (neither acidic nor basic). The nucleus has a distinct lobed appearance and may have two to five lobes, the number increasing with the age of the cell.
Neutrophils are rapid responders to the site of infection and are efficient phagocytes with a preference for bacteria. Their granules include lysozyme, an enzyme capable of lysing, or breaking down, bacterial cell walls and defensins, proteins that bind to and puncture bacterial and fungal plasma membranes causing the cell contents to leak out. Abnormally high counts of neutrophils indicate infection and/or inflammation, particularly triggered by bacteria, but are also found in burn patients and others experiencing unusual stress. A burn injury increases the proliferation of neutrophils in order to fight off infection that can result from the destruction of the barrier of the skin. Low counts may be caused by drug toxicity and other disorders, and may increase an individual’s susceptibility to infection.
Eosinophils typically represent 2–4 percent of total leukocyte count. The granules of eosinophils stain best with an acidic stain known as eosin. The granules of eosinophils include antihistamine molecules, which counteract the activities of histamines, inflammatory chemicals produced by basophils and other inflammatory cells. Some eosinophil granules contain molecules toxic to parasitic worms, which can enter the body either through the integument or when an individual consumes raw or undercooked fish or meat. Eosinophils are also capable of phagocytosis. High counts of eosinophils are typical of patients experiencing allergies, parasitic worm infestations, and some autoimmune diseases. Low counts may be due to drug toxicity and stress.
Basophils are the least common leukocyte, typically comprising less than one percent of the total leukocyte count. The granules of basophils stain best with basic (alkaline) stains. In general, basophils intensify the inflammatory response. The granules of basophils release histamines, which contribute to inflammation, and heparin, which opposes blood clotting. High counts of basophils are associated with allergies, parasitic infections, and hypothyroidism. Low counts are associated with pregnancy, stress, and hyperthyroidism.
Agranular Leukocytes: Agranular leukocytes contain smaller, less-visible granules in their cytoplasm than do granular leukocytes. The nucleus is simple in shape, sometimes with an indentation but without distinct lobes. There are two major types of agranulocytes: lymphocytes and monocytes.
Lymphocytes are the primary cells of adaptive immune responses (Table 1). They are the only formed element of blood that arises from lymphoid stem cells. Although they initially form in the bone marrow, much of their subsequent development and reproduction occurs in the lymphatic tissues. Lymphocytes are the second most common type of leukocyte, accounting for about 20–30 percent of all leukocytes, and are essential for the immune response.
Table 1: Lymphocytes
| Type of Lymphocyte | Primary Function |
|---|---|
| B Lymphocyte | Generates diverse antibodies
Memory for subsequent infections |
| T Lymphocyte | Secretes chemical messengers
Cytotoxic activity Memory for subsequent infections |
| Natural Killer Cell | Destroys virally infected cells |
Abnormally high lymphocyte counts are characteristic of viral infections as well as some types of cancer. Abnormally low lymphocyte counts are characteristic of prolonged (chronic) illness or immunosuppression, including that caused by HIV infection and drug therapies that often involve steroids.
The two basic types of lymphocytes, B cells and T cells (also called B lymphocytes and T lymphocytes), are identical morphologically, with a large, often spherical, central nucleus surrounded by a thin layer of cytoplasm. They are distinguished from each other by their surface protein markers as well as by the molecules they secrete. B cells mature in red bone marrow and T cells mature in the thymus. B cells and T cells are found in many parts of the body, circulating in the bloodstream and lymph, and residing in secondary lymphoid organs, including the spleen and lymph nodes. The human body contains approximately 1012 lymphocytes. Both B cells and T cells play prominent roles in defending the body against specific pathogens (disease-causing microorganisms) and are involved in specific immunity.
One form of B cells, when activated, become plasma cells. These cells differ in morphology from standard B and T cells in that they contain a large amount of cytoplasm packed with the protein-synthesizing machinery known as rough endoplasmic reticulum. A plasma cell forms from a naïve B cell with the purpose of producing antibodies or immunoglobulins. An antibody is any of the group of proteins that binds specifically to pathogen-associated molecules known as antigens. An antigen is a chemical structure on the surface of a pathogen, or the soluble product of a pathogen (ie. a toxin), that binds to T or B lymphocyte receptors. Once activated by binding to antigen, B cells differentiate into plasma cells and begin producing and secreting large quantities of antigen specific antibodies. These travel through the body targeting pathogens or toxins for destruction using mechanisms that will be discussed later in the chapter. This is also referred to as humoral (body fluid) immunity.
The T cell, on the other hand, does not secrete antibody but performs a variety of functions in the adaptive immune response. Different T cell types have the ability to either secrete soluble factors that communicate with and activate other cells of the adaptive immune response or destroy cells infected with intracellular pathogens. Therefore, T cells provide cell-mediated immunity by physically attacking foreign or diseased cells. Both B and T cells can differentiate to memory cells that form after exposure to a pathogen and mount rapid responses upon subsequent exposures. Unlike other leukocytes, memory cells live for many years. The roles of T and B lymphocytes in the adaptive immune response will be discussed further on.
Another important lymphocyte is the natural killer cell, a participant in the innate immune response. A natural killer (NK) cell is a circulating blood cell that contains cytotoxic (cell-killing) granules in its extensive cytoplasm. It shares this mechanism with the cytotoxic T cells of the adaptive immune response. NK cells are capable of recognizing cells that do not express “self” proteins on their plasma membrane or that contain foreign or abnormal markers. These “non-self” cells include cancer cells, cells infected with a virus, and other cells with atypical surface proteins. Thus, they provide generalized, nonspecific immunity and are among the body’s first lines of defense against viruses and certain types of cancer.
Monocytes originate from myeloid stem cells. They normally represent 2–8 percent of the total leukocyte count. Macrophages are monocytes that have left the circulation and phagocytize debris, foreign pathogens, and many dead, worn out, or damaged cells, including red blood cells. Macrophages also release antimicrobial defensins and chemotactic chemicals that attract other leukocytes to the site of an infection. Some macrophages occupy fixed locations, whereas others wander through the tissue fluid.
Abnormally high counts of monocytes are associated with certain viral or fungal infections, tuberculosis, and some forms of leukemia and other chronic diseases. Abnormally low counts are typically caused by suppression of the bone marrow due to drugs or infiltration by tumor cells.
Part 2: The Lymphatic and Immune System
In June 1981, the Centers for Disease Control and Prevention (CDC), in Atlanta, Georgia, published a report of an unusual cluster of five patients in Los Angeles, California. All five were diagnosed with a rare pneumonia caused by a fungus called Pneumocystis jirovecii (formerly known as Pneumocystis carinii).
Why was this unusual? Although commonly found in the lungs of healthy individuals, this fungus is an opportunistic pathogen that causes disease in individuals with suppressed or underdeveloped immune systems. The very young, whose immune systems have yet to mature, and the elderly, whose immune systems have declined with age, are particularly susceptible. The five patients from LA, though, were between 29 and 36 years of age and should have been in the prime of their lives, immunologically speaking. What could be going on?
A few days later, a cluster of eight cases was reported in New York City, also involving young patients, this time exhibiting a rare form of skin cancer known as Kaposi’s sarcoma. This cancer of the cells that line the blood and lymphatic vessels was previously observed as a relatively innocuous disease of the elderly. The disease that doctors saw in 1981 was frighteningly more severe, with multiple, fast-growing lesions that spread to all parts of the body, including the trunk and face. Could the immune systems of these young patients have been compromised in some way? Indeed, when they were tested, they exhibited extremely low numbers of a specific type of white blood cell in their bloodstreams, indicating that they had somehow lost a major part of the immune system.
Acquired immune deficiency syndrome, or AIDS, turned out to be a new disease caused by the previously unknown human immunodeficiency virus (HIV). Although nearly 100 percent fatal in those with active HIV infections in the early years, the development of anti-HIV drugs has transformed HIV infection into a chronic, manageable disease and not the certain death sentence it once was. One positive outcome resulting from the emergence of HIV disease was that the public’s attention became focused as never before on the importance of having a functional and healthy immune system.
Anatomy of the Lymphatic and Immune Systems: The immune system is the complex collection of cells and organs that destroys or neutralizes pathogens that would otherwise cause disease or death. The lymphatic system is the system of vessels, cells, and organs that carries excess fluids to the bloodstream and filters pathogens from the blood. The swelling of lymph nodes during an infection and the transport of lymphocytes via the lymphatic vessels are but two examples of the many connections between these critical organ systems.
Functions of the Lymphatic System: A major function of the lymphatic system is to drain body fluids and return them to the bloodstream. Blood pressure causes leakage of fluid from the capillaries, resulting in the accumulation of fluid in the interstitial space—that is, spaces between individual cells in the tissues. In humans, 20 liters of plasma is released into the interstitial space of the tissues each day due to capillary filtration. Once this filtrate is out of the bloodstream and in the tissue spaces, it is referred to as interstitial fluid. Of this, 17 liters is reabsorbed directly by the blood vessels. But what happens to the remaining three liters? This is where the lymphatic system comes into play. It drains the excess fluid and empties it back into the bloodstream via a series of vessels, trunks, and ducts. Lymph is the term used to describe interstitial fluid once it has entered the lymphatic system. When the lymphatic system is damaged in some way, such as by being blocked by cancer cells or destroyed by injury, protein-rich interstitial fluid accumulates (sometimes “backs up” from the lymph vessels) in the tissue spaces. This inappropriate accumulation of fluid referred to as lymphedema may lead to serious medical consequences.
As the vertebrate immune system evolved, the network of lymphatic vessels became convenient avenues for transporting the cells of the immune system. Additionally, dietary lipids and fat-soluble vitamins absorbed in the gut use this system of transport.
Cells of the immune system not only use lymphatic vessels to make their way from interstitial spaces back into the circulation, but they also use lymph nodes as major staging areas for the development of critical immune responses. A lymph node is one of the small, bean-shaped organs located throughout the lymphatic system.
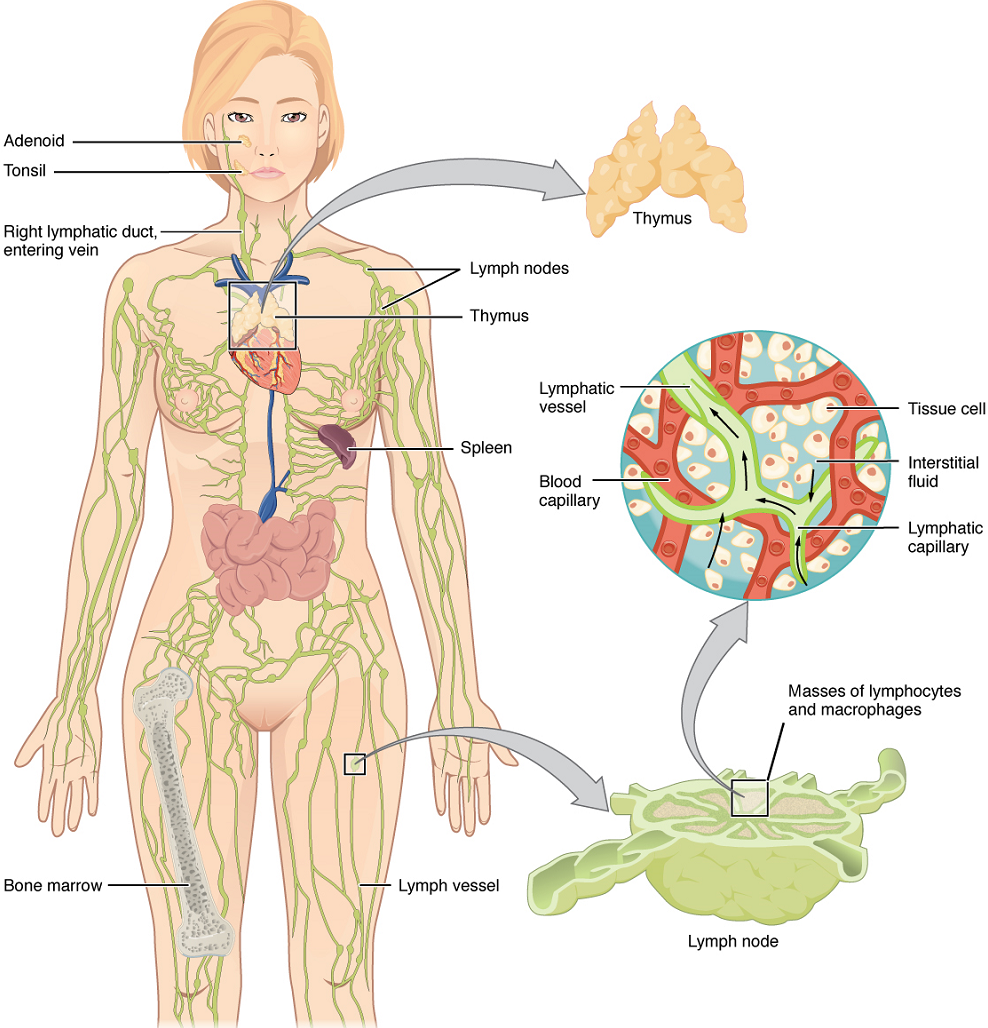
Structure of the Lymphatic System: The lymphatic vessels begin as open-ended capillaries, which feed into larger and larger lymphatic vessels, and eventually empty into the bloodstream by a series of ducts. Along the way, the lymph travels through the lymph nodes, which are commonly found near the groin, armpits, neck, chest, and abdomen. Humans have about 500–600 lymph nodes throughout the body (Figure 3).
A major distinction between the lymphatic and cardiovascular systems in humans is that lymph is not actively pumped by the heart, but is forced through the vessels by the movements of the body, the contraction of skeletal muscles during body movements, and breathing. One-way valves (semi-lunar valves) in lymphatic vessels keep the lymph moving toward the heart. Lymph flows from the lymphatic capillaries, through lymphatic vessels, and then re-enters the circulatory system via the lymphatic ducts located at the junction of the jugular and subclavian veins in the neck.
Lymphatic Capillaries: Lymphatic capillaries, also called the terminal lymphatics, are vessels where interstitial fluid enters the lymphatic system to become lymph fluid. Located in almost every tissue in the body, these vessels are interlaced among the arterioles and venules of the circulatory system in the soft connective tissues of the body (Figure 4). Exceptions are the central nervous system, bone marrow, bones, teeth, and the cornea of the eye, which do not contain lymph vessels.
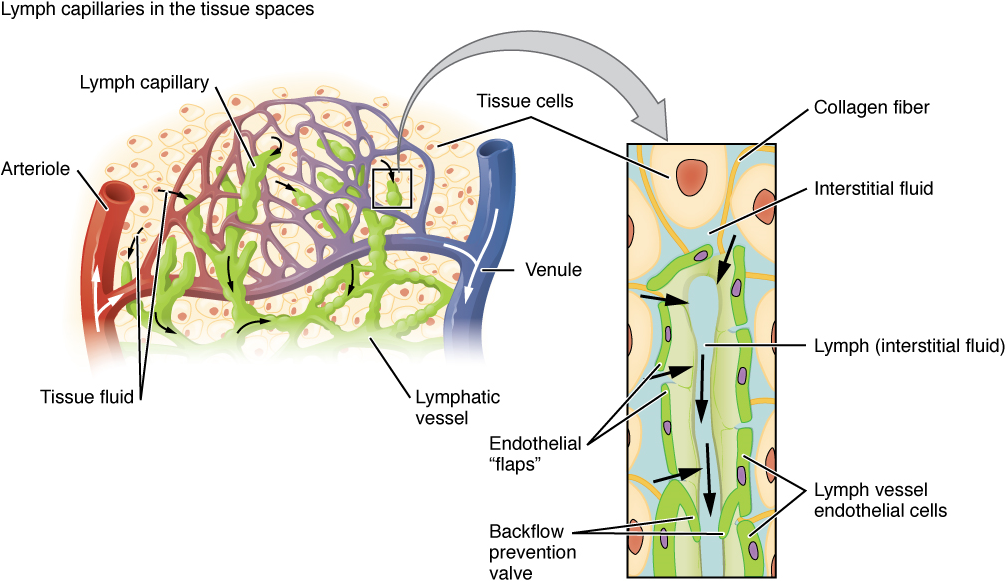
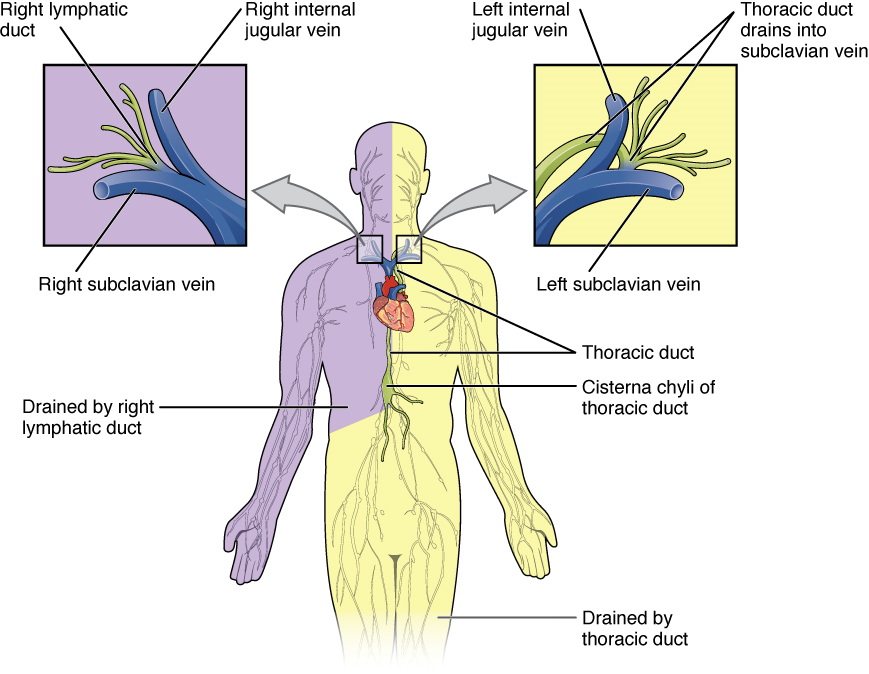
Larger Lymphatic Vessels, Trunks, and Ducts: The lymphatic capillaries empty into larger lymphatic vessels, which are similar to veins in terms of their three-tunic structure and the presence of valves. These one-way valves are located fairly close to one another, and each one causes a bulge in the lymphatic vessel, giving the vessels a beaded appearance (see Figure 4). The superficial and deep lymphatics eventually merge to form larger lymphatic vessels known as lymphatic trunks. On the right side of the body, the right sides of the head, thorax, and right upper limb drain lymph fluid into the right subclavian vein via the right lymphatic duct (Figure 5). On the left side of the body, the remaining portions of the body drain into the larger thoracic duct, which drains into the left subclavian vein.
Primary Lymphoid Organs and Lymphocyte Development: Understanding the differentiation and development of B and T cells is critical to the understanding of the adaptive immune response. It is through this process that the body (ideally) learns to destroy only pathogens and leaves the body’s own cells relatively intact. The primary lymphoid organs are the bone marrow and thymus gland. The lymphoid organs are where lymphocytes mature, proliferate, and are selected, which enables them to attack pathogens without harming the cells of the body.
Bone Marrow: The bone marrow is responsible for most hematopoietic functions, although the final stages of the differentiation of some cells may take place in other organs. The red bone marrow is a loose collection of cells where hematopoiesis occurs, and the yellow bone marrow is a site of energy storage, which consists largely of fat cells (Figure 6). The B cell undergoes nearly all of its development in the red bone marrow, whereas the immature T cell, called a thymocyte, leaves the bone marrow and matures largely in the thymus gland.
Thymus: The thymus gland is a bilobed organ found in the space behind the sternum and anterior to the heart, it overlies the aortic arch, superior vena cava and trachea. The organ contains large numbers of thymocytes with some epithelial cells, macrophages, and dendritic cells (two types of phagocytic cells that are derived from monocytes). As mentioned, thymocytes mature into T cells in the thymus.
Secondary Lymphoid Organs and their Roles in Active Immune Responses: Lymphocytes develop and mature in the primary lymphoid organs, but they mount immune responses from the secondary lymphoid organs. A naïve lymphocyte is one that has left the primary organ and entered a secondary lymphoid organ. Naïve lymphocytes are fully functional immunologically, but have yet to encounter an antigen to respond to. In addition to circulating in the blood and lymph, lymphocytes concentrate in secondary lymphoid organs, which include the lymph nodes, spleen, and lymphoid nodules such as the tonsils.
Lymph Nodes: Lymph nodes function to remove debris and pathogens from the lymph, and are thus sometimes referred to as the “filters of the lymph”. Any bacteria that infect the interstitial fluid are taken up by the lymphatic capillaries and transported to a regional lymph node. Dendritic cells and macrophages within this organ internalize and kill many of the pathogens that pass through, thereby removing them from the body. The lymph node is also the site of adaptive immune responses mediated by T cells, B cells, and accessory cells of the adaptive immune system.
Spleen: In addition to the lymph nodes, the spleen is a major secondary lymphoid organ. It is about 12 cm (5 in) long and is attached to the lateral border of the stomach via the gastrosplenic ligament. The spleen is sometimes called the “filter of the blood” because of its extensive vascularization and the presence of macrophages and dendritic cells that remove microbes and other materials from the blood, including dying red blood cells. The spleen also functions as the location of immune responses to blood-borne pathogens.
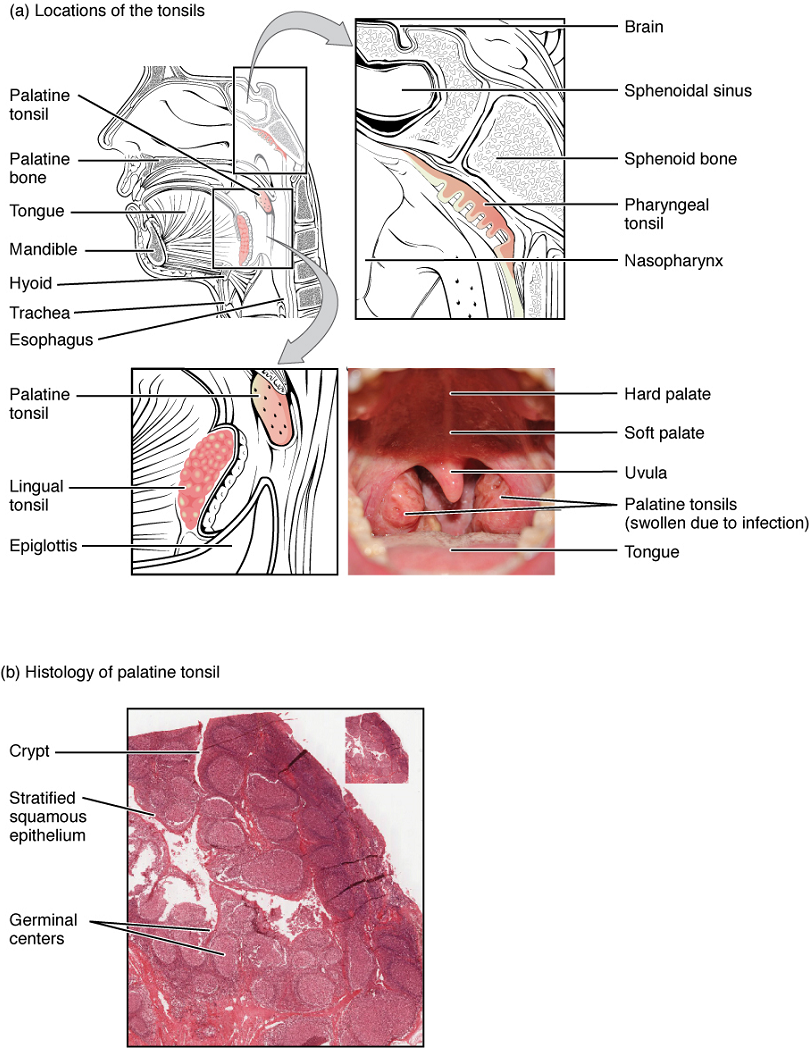
Tonsils: These lymphoid nodules located along the inner surface of the pharynx are important in developing immunity to oral pathogens (Figure 7). The tonsil located at the back of the throat, called the pharyngeal tonsil, is sometimes referred to as the adenoid when swollen. Such swelling is an indication of an active immune response to infection. This seems to be the major function of tonsils—to help children’s bodies recognize, destroy, and develop immunity to common environmental pathogens so that they will be protected in their later lives. Tonsils are often removed in those children who have recurring throat infections, especially those involving the palatine tonsils on either side of the throat, whose swelling may interfere with their breathing and/or swallowing.

Watch this CrashCourse video to learn more about the lymphatic system! Direct link: https://youtu.be/I7orwMgTQ5I
The Organization of Immune Function: The immune system is a collection of barriers, cells, and soluble proteins that interact and communicate with each other in extraordinarily complex ways. The modern model of immune function is organized into three phases based on the timing of their effects. The three temporal phases consist of the following:
- Barrier defenses such as the skin and mucous membranes, which act instantaneously to prevent pathogenic invasion into the body tissues.
- The rapid but nonspecific innate immune response, which consists of a variety of specialized cells and soluble factors.
- The slower but more specific and effective adaptive immune response, which involves many cell types and soluble factors, but is primarily controlled by white blood cells (leukocytes) known as lymphocytes, which help control immune responses.
Barrier Defenses and the Innate Immune Response: The immune system can be divided into two overlapping mechanisms to destroy pathogens: the innate immune response, which is relatively rapid but nonspecific and thus not always effective, and the adaptive immune response, which is slower in its development following infection, but is highly specific and effective at attacking a wide variety of pathogens (Figure 8).
Any discussion of the innate immune response usually begins with the physical barriers that prevent pathogens from entering the body, destroy them after they enter, or flush them out before they can establish themselves in the hospitable environment of the body’s soft tissues. Barrier defenses are part of the body’s most basic defense mechanisms. The barrier defenses are not a response to infections, but they are continuously working to protect against a broad range of pathogens.
The different modes of barrier defenses are associated with the external surfaces of the body, where pathogens may try to enter (Table 2). The primary barrier to the entrance of microorganisms into the body is the skin. Not only is the skin covered with a layer of dead, keratinized epithelium that is too dry for bacteria in which to grow, but as these cells are continuously sloughed off from the skin, they carry bacteria and other pathogens with them. Additionally, sweat and other skin secretions may lower pH, contain toxic lipids, contain antimicrobial peptides such as dermcidin, and physically wash microbes away.
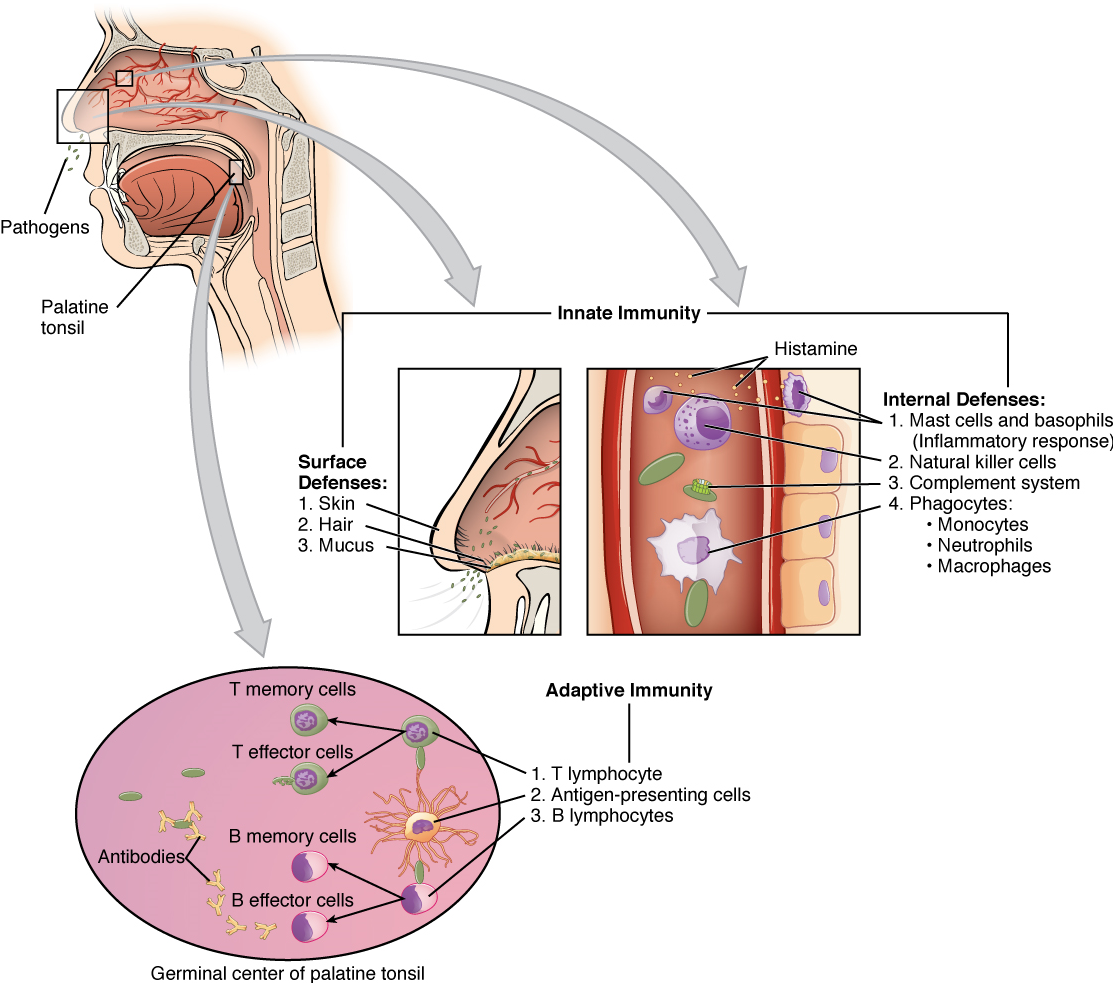
Another barrier is the saliva in the mouth, which is rich in lysozyme—an enzyme that destroys bacteria by digesting their cell walls. The acidic environment of the stomach, which is fatal to many pathogens, is also a barrier. Additionally, the mucus layer of the gastrointestinal tract, respiratory tract, reproductive tract, eyes, ears, and nose traps both microbes and debris, and facilitates their removal. In the case of the upper respiratory tract, ciliated epithelial cells move potentially contaminated mucus upwards to the mouth, where it is then swallowed into the digestive tract, ending up in the harsh acidic environment of the stomach. Considering how often you breathe compared to how often you eat or perform other activities that expose you to pathogens, it is not surprising that multiple barrier mechanisms have evolved to work in concert to protect this vital area.
Cells of the Innate Immune Response: A phagocyte is a cell that is able to surround and engulf a particle or cell, a process called phagocytosis. The phagocytes of the immune system engulf other particles or cells, either to clean an area of debris, remove old cells, or to kill pathogenic organisms such as bacteria. The phagocytes are the body’s fast acting, first line of immunological defense against organisms that have breached barrier defenses and have entered the vulnerable tissues of the body.
| Site | Defensive structure | Protective aspect |
|---|---|---|
| Skin (physical structure) | Epidermal surface | Keratinized cells of surface, Langerhans cells |
| Skin (secretions) | Eccrine glands | Low pH, dermcidin, washing action |
| Oral cavity | Salivary glands | Lysozyme |
| Stomach | Gastric juice | Low pH |
| Mucous membranes | Mucosal epithelium | Layered cells |
| Mucous membranes (secretions) | Cells producing mucus | Traps pathogens, dust, debris, etc.; washing action; defensins and lysozyme |
| Skin and mucosal surfaces | Normal flora (nonpathogenic bacteria) | Compete with pathogenic microbes |
Phagocytes: Macrophages and Neutrophils:
Many of the cells of the immune system have a phagocytic ability, at least at some point during their life cycles. Phagocytosis is an important and effective mechanism of destroying pathogens during innate immune responses. The phagocyte takes the organism inside itself as a phagosome, which subsequently fuses with a lysosome and its digestive enzymes, forming a phagolysosome, and thus effectively killing many pathogens. On the other hand, some bacteria including Mycobacterium tuberculosis, the pathogen causing tuberculosis, may be resistant to these enzymes and are therefore much more difficult to clear from the body. Macrophages, neutrophils, and dendritic cells are the major phagocytes of the immune system.
A macrophage is an irregularly shaped phagocyte that is amoeboid in nature and is the most versatile of the phagocytes in the body. Macrophages move through tissues and squeeze through capillary walls using pseudopodia. They not only participate in innate immune responses but have also evolved to cooperate with lymphocytes as part of the adaptive immune response. Macrophages exist in many tissues of the body, either freely roaming through connective tissues or fixed to reticular fibers within specific tissues such as lymph nodes. When pathogens breach the body’s barrier defenses, macrophages are the first line of defense (Table 3). They are called different names, depending on the tissue: Kupffer cells in the liver, histiocytes in connective tissue, microglia in the brain, and alveolar macrophages in the lungs.
A neutrophil is a phagocytic cell that is attracted via chemotaxis from the bloodstream to infected tissues. Whereas macrophages act like sentries, always on guard against infection, neutrophils can be thought of as military reinforcements that are called into a battle to hasten the destruction of the enemy. Neutrophils are usually thought of as the primary pathogen-killing cell of the inflammatory process of the innate immune response.
A monocyte is a circulating precursor cell that differentiates into either a macrophage or dendritic cell. Monocytes can be rapidly attracted to areas of infection by signal molecules of inflammation.
| Cell | Cell type | Primary location | Function in the innate immune response |
|---|---|---|---|
| Macrophage | Agranulocyte | Body cavities/organs | Phagocytosis |
| Dendritic cell | Agranulocyte | Skin and mucous membranes | Phagocytosis |
| Neutrophil | Granulocyte | Blood | Phagocytosis |
| Monocyte | Agranulocyte | Blood | Precursor of macrophages and dendritic cells |
Natural Killer Cells: NK cells, as mentioned previously, are a type of lymphocyte that play an important role in the innate immune response. They have the ability to induce apoptosis, that is, programmed cell death, in cells infected with intracellular pathogens such as obligate intracellular bacteria (for example, Mycobacteria) and viruses. NK cells recognize these cells by mechanisms that are still not well understood, but that presumably involve their surface receptors. NK cells can induce apoptosis, in which a cascade of events inside the cell causes its own death. In addition, NK cells secrete chemicals which enhance inflammation.
Should the cells of the innate immune system come into contact with a species of pathogen they recognize, the cell will bind to the pathogen and initiate phagocytosis (or cellular apoptosis in the case of an intracellular pathogen) in an effort to destroy the offending microbe.
Soluble Mediators of the Innate Immune Response: These are soluble factors secreted during innate or early induced responses, and later during adaptive immune responses. Examples include signaling molecules such as cytokines or chemokines used to recruit and activate immunological cells. Proteins involved in the complement system are also important mediators of the immune response. The complement system is a series of signaling cascades with functions such as labeling pathogens for phagocytosis (opsonization) or killing pathogens by directly damaging the plasma membrane.
Inflammatory Response: Everyone has experienced inflammation at some point in their lives. Stub a toe, cut a finger, or perform any activity that causes tissue damage and inflammation will result, with its four characteristics: heat, redness, pain, and swelling (“loss of function” is sometimes mentioned as a fifth characteristic). It is important to note that inflammation does not have to be initiated by an infection, but can also be caused by tissue injuries. The release of damaged cellular contents into the site of injury is enough to stimulate the response, even in the absence of breaks in physical barriers that would allow pathogens to enter (by hitting your thumb with a hammer, for example). The inflammatory reaction brings in phagocytic cells to the damaged area to clear cellular debris and to set the stage for wound repair (Figure 9).
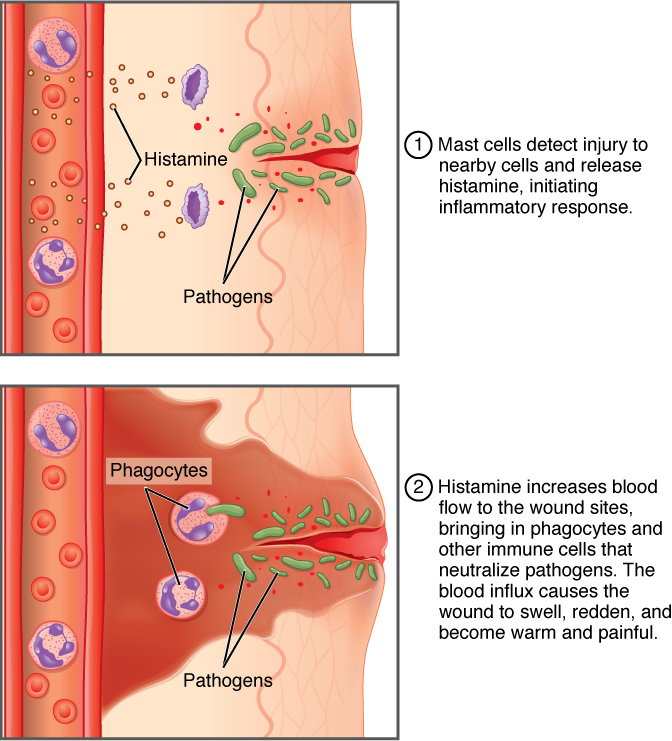
This reaction also brings in the cells of the innate immune system, allowing them to get rid of the sources of a possible infection or injury. Inflammation is part of a very basic form of immune response. The process not only brings fluid and cells into the site of damage to destroy the pathogen and remove it and any debris from the site, but it also helps to isolate the site, limiting the spread of the pathogen. Acute inflammation is a short-term inflammatory response to an insult to the body. If the cause of the inflammation is not resolved, however, it can lead to chronic inflammation, which is associated with major tissue destruction and fibrosis. Chronic inflammation is ongoing inflammation. It can be caused by foreign bodies, persistent pathogens, and autoimmune diseases such as rheumatoid arthritis.
There are four important parts to the inflammatory response:
- Tissue Injury. The released contents of injured cells stimulate the release of mast cell granules and their potent inflammatory mediators. These mediators are involved in vasodilation, increasing permeability of local capillaries, and recruitment of phagocytes.
- Vasodilation. Many inflammatory mediators such as histamine are vasodilators that increase the diameters of local capillaries. This causes increased blood flow and is responsible for the heat and redness of inflamed tissue. It allows greater access of the immune components of blood to the site of inflammation.
- Increased Vascular Permeability. At the same time, inflammatory mediators increase the permeability of the local vasculature, causing leakage of fluid into the interstitial space, resulting in the swelling, or edema, associated with inflammation. This allows immune cells and mediators to exit the blood stream and enter the site of infection or injury.
- Recruitment of Phagocytes. Inflammatory mediators also attract neutrophils from the blood to the site of infection by chemotaxis. Following an early neutrophil infiltration stimulated by macrophage signals, more macrophages are recruited to clean up the debris remaining at the site. When local infections are severe, neutrophils are attracted to the sites of infections in large numbers, and as they phagocytose the pathogens and subsequently die, their accumulated cellular remains are visible as pus at the infection site.
Overall, inflammation is valuable for many reasons. Not only are the pathogens killed and debris removed, but the increase in vascular permeability encourages the entry of clotting factors, the first step towards wound repair. Inflammation also facilitates the transport of antigen to lymph nodes by macrophages or dendritic cells for the development of the adaptive immune response.
Fever: The mechanisms of inflammation described so far are primarily local. Another inflammatory response that is systemic in nature is that of fever. Fever is defined as an increase in the set-point of the body’s thermostat, with the result that homeostatic mechanisms raise the temperature of the body above the normal of about 37ºC.
The increase in temperature has several effects that are beneficial to the body’s defense. These include increasing the activity of the immune system (e.g., enhancing the efficiency of white blood cells). Fever also results in an increase in the production of the iron-binding protein transferrin which reduces the availability of iron in the blood, which in turn can reduce the rate of growth of microbes. The beneficial effects of such an increase in body temperature disappear, however, should the value go over 41ºC, as human proteins begin denaturing.

The Adaptive Immune Response
T lymphocytes and their Functional Types: Innate immune responses (and early induced responses) are in many cases ineffective at completely controlling pathogen growth. However, they slow pathogen growth and allow time for the adaptive immune response to strengthen, and either control or eliminate the pathogen. The innate immune system also sends signals to the cells of the adaptive immune system, guiding them in how to attack the pathogen. Thus, the innate and adaptive mechanisms are two important arms of the immune response.
The Benefits of the Adaptive Immune Response: The specificity of the adaptive immune response—its ability to specifically recognize and make a response against a wide variety of pathogens—is its great strength. Antigens, the small chemical groups often associated with pathogens and their products, are recognized by receptors on the surfaces of B and T lymphocytes. The adaptive immune response to these antigens is so versatile that it can respond to nearly any pathogen. This increase in specificity comes because the adaptive immune response has a unique way to develop as many as 1011, or 100 trillion, different receptors to recognize nearly every conceivable pathogen. Immunological memory is another benefit of adaptive immunity and this will be described further.
Primary Disease and Immunological Memory: The immune system’s first exposure to a pathogen is called a primary adaptive response. Symptoms of a first infection, called primary disease, are always relatively severe because it takes time for an initial adaptive immune response to a pathogen to become effective.
Upon re-exposure to the same pathogen, a secondary adaptive immune response is generated, which is stronger and faster that the primary response. The secondary adaptive response often eliminates a pathogen before it can cause significant tissue damage or any symptoms. Without symptoms, there is no disease, and the individual is not even aware of the infection. This secondary response is the basis of immunological memory, which protects us from getting diseases repeatedly from the same pathogen. By this mechanism, an individual’s exposure to pathogens early in life spares the person from these diseases later in life.
Self-Recognition: Another important feature of the adaptive immune response is its ability to distinguish between self-antigens, those that are normally present in the body, and foreign antigens, those that might be on a potential pathogen. As T and B cells mature, there are mechanisms in place that prevent them from recognizing self-antigen, preventing a damaging immune response against the body. These mechanisms are not 100 percent effective, however, and their breakdown leads to autoimmune diseases, which will be discussed later.
T Cell-Mediated Immune Responses: The primary cells that control the adaptive immune response are the lymphocytes, the T and B cells. T cells are particularly important, as they not only control a multitude of immune responses directly, but also control B cell immune responses in many cases as well. Thus, many of the decisions about how to attack a pathogen are made at the T cell level, and knowledge of their functional types is crucial to understanding the functioning and regulation of adaptive immune responses as a whole.
Antigens: Antigens on pathogens are usually large and complex, and can be either carbohydrate or protein based. Each T cell produces only one type of receptor and thus is specific for a single particular antigen. It is the interaction of the shape of the antigen and the complementary shape of the antigen-binding receptor that accounts for the chemical basis of specificity.
Antigen-presenting Cells: Antigen-presenting cells represent an important link between the innate and adaptive immune response. These stimulators of the adaptive response include macrophages, dendritic cells, and B cells. Macrophages stimulate T cells to release cytokines that enhance phagocytosis by macrophages. Dendritic cells also kill pathogens by phagocytosis, but both have the additional function of bringing antigens to regional draining lymph nodes. The lymph nodes are the locations in which most T cell responses against pathogens of the interstitial tissues are mounted. B cells may also present antigens to T cells, which are necessary for certain types of antibody responses, to be covered later in this chapter.
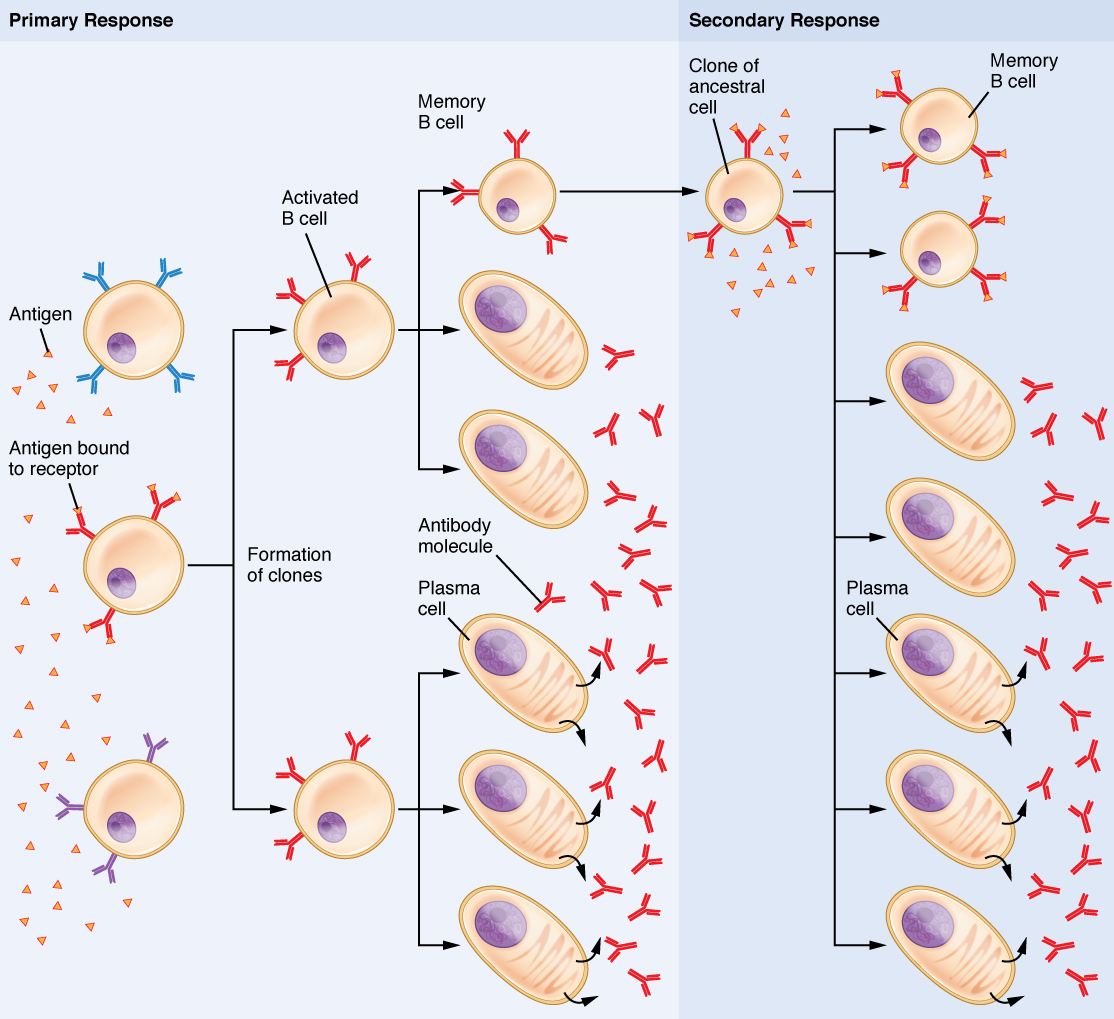
Mechanisms of T Cell-mediated Immune Responses: Mature T cells become activated by recognizing foreign antigen on an antigen presenting cell and begin dividing rapidly by mitosis. This proliferation of T cells is called clonal expansion and is necessary to make the immune response strong enough to effectively control a pathogen. Only those clones of lymphocytes whose receptors are activated by the antigen are stimulated to proliferate. Once activated, the selected clones increase in number and make many copies of each cell type, each clone with its unique receptor. By the time this process is complete, the body will have large numbers of specific lymphocytes available to fight the infection (see Figure 10).
The Cellular Basis of Immunological Memory: As already discussed, one of the major features of an adaptive immune response is the development of immunological memory.
During a primary adaptive immune response, both memory T cells and effector T cells are generated. Memory T cells are long-lived and can even persist for a lifetime. Memory cells are primed to act rapidly. Thus, any subsequent exposure to the pathogen will elicit a very rapid T cell response. This rapid, secondary adaptive response generates large numbers of effector T cells so fast that the pathogen is often overwhelmed before it can cause any symptoms of disease. This is what is meant by immunity to a disease. The same pattern of primary and secondary immune responses occurs in B cells and the antibody response, as will be discussed later in the chapter.
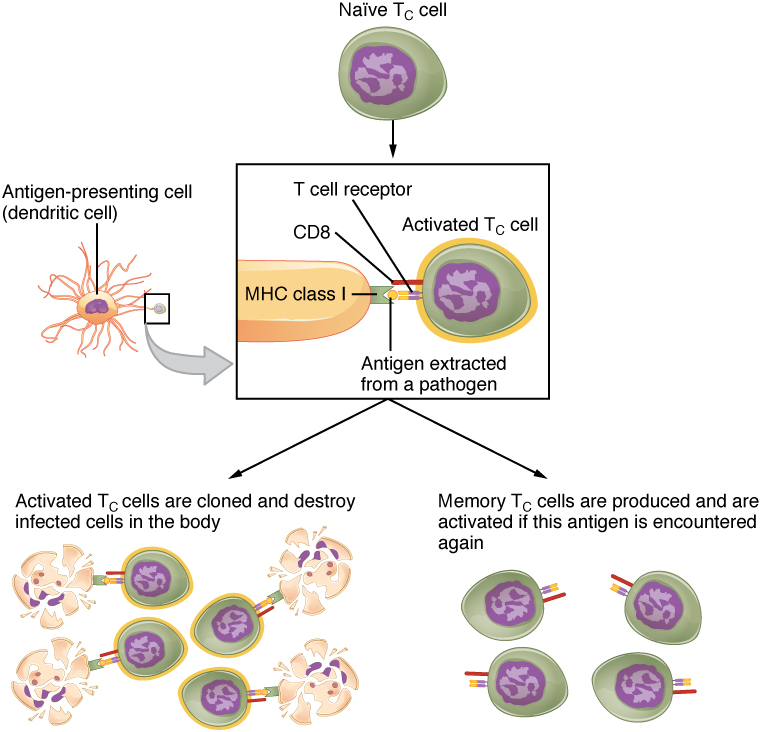
T Cell Types and their Functions: T cells can contain cell adhesion molecules that keep the T cell in close contact with the antigen-presenting cell by directly binding to the antigen-presenting receptor on its membrane. These markers are either CD4 or CD8 molecules (CD refers to Cluster of Differentiation) (Figure 11).
Although the correlation is not absolute, CD4-bearing T cells are associated with helper functions and CD8-bearing T cells are associated with cytotoxicity. These functional distinctions based on CD4 and CD8 markers are useful in defining the function of each type.
Helper T Cells and their Cytokines: Helper T cells (Th), bearing the CD4 molecule, function by secreting cytokines that act to enhance other immune responses. There are two classes of Th cells, and they act on different components of the immune response. These cells are not distinguished by their surface molecules but by the characteristic set of cytokines they secrete.
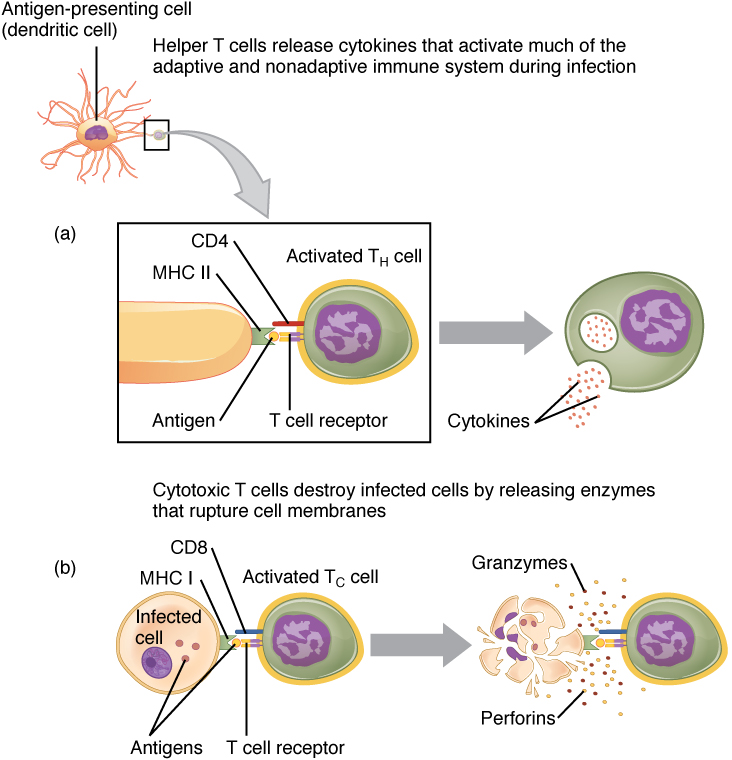
Th1 cells are a type of helper T cell that secretes cytokines that regulate the immunological activity and development of a variety of cells, including macrophages and other types of T cells.
Th2 cells, on the other hand, are cytokine-secreting cells that act on B cells to drive their differentiation into plasma cells that make antibody.
Cytotoxic T cells: Cytotoxic T cells (Tc) are T cells that kill target specific cells by inducing apoptosis using the same mechanism as NK cells. In addition, as long as the antigen is recognized by the cell, each Tc cell can kill more than one target cell, making them especially effective. While they are active in all pathogenic infections, Tc cells are so important in the antiviral immune response that some speculate that this was the main reason the adaptive immune response evolved in the first place.
B-lymphocytes and Antibodies: Antibodies were the first component of the adaptive immune response to be characterized by scientists working on the immune system. It was already known that individuals who survived a bacterial infection were immune to re-infection with the same pathogen. Early microbiologists took serum from an immune patient and mixed it with a fresh culture of the same type of bacteria, then observed the bacteria under a microscope. The bacteria became clumped in a process called agglutination. When a different bacterial species was used, the agglutination did not happen. Thus, there was something in the serum of immune individuals that could specifically bind to and agglutinate bacteria. Scientists now know the cause of the agglutination is an antibody molecule, also called an immunoglobulin.
What is an antibody? An antibody protein is essentially a secreted form of a B cell receptor. (In fact, surface immunoglobulin is another name for the B cell receptor.) Not surprisingly, the same genes encode both the secreted antibodies and the surface immunoglobulins. One minor difference in the way these proteins are synthesized distinguishes a naïve B cell with antibody on its surface from an antibody-secreting plasma cell with no antibodies on its surface. The antibodies of the plasma cell have the exact same antigen-binding site and specificity as their B cell precursors.
There are five different classes of antibody (also called immunoglobulin Ig) found in humans: IgM, IgD, IgG, IgA, and IgE. Each of these has specific functions in the immune response, so by learning about them, researchers can learn about the great variety of antibody functions critical to many adaptive immune responses.
B Cell Differentiation and Activation: B cells differentiate in the bone marrow. During the process of maturation, up to 100 trillion different clones of B cells are generated, which is similar to the diversity of antigen receptors seen in T cells.
After B cells are activated by their binding to antigen, they differentiate into plasma cells. Plasma cells often leave the secondary lymphoid organs, where the response is generated, and migrate back to the bone marrow, where the whole differentiation process from a lymphoid progenitor cell started. After secreting antibodies for a specific period, plasma cells die, as most of their energy is devoted to making antibodies and not to maintaining themselves. Thus, plasma cells are said to be terminally differentiated.
The final B cell of interest is the memory B cell, which results from the clonal expansion of an activated B cell. Memory B cells function in a way similar to memory T cells. They lead to a stronger and faster secondary response when compared to the primary response, as illustrated below.
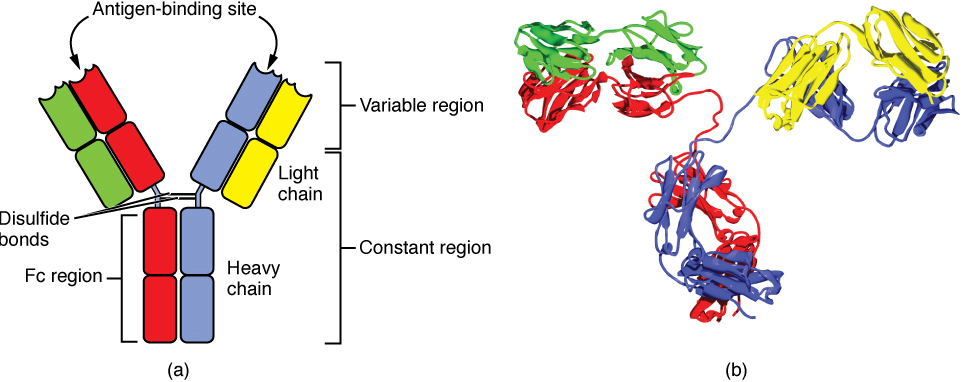
Antibody Structure: Antibodies are glycoproteins consisting of two types of polypeptide chains with attached carbohydrates. The heavy chain and the light chain are the two polypeptides that form the antibody, two of each are required to form a generic antibody structure. The main differences between the classes of antibodies are in the differences between their heavy chains, but as you shall see, the light chains have an important role, forming part of the antigen-binding site on the antibody molecules (Figure 12).
Functions of Antibodies: In general, antibodies have two basic functions. They can act as the B cell antigen receptor or they can be secreted, circulate, and bind to a pathogen, often labeling it for identification by other forms of the immune response.
Effects of Antibody-Antigen Binding: Antibodies that bind to antigens can lead to a number of different outcomes, depending on the nature of the antigen and the structure of the antibody. In a process called neutralization, antibodies bind to antigens on the surface of some viruses, or to toxins secreted by bacteria, in a way that prevents them from negatively affecting body cells. The antibodies neutralize the pathogen or toxin by physically covering up the dangerous parts so it cannot damage body cells. Antibodies have at least two antigen-binding sites and therefore they can bind to antigen on the surface of two or more cells or to multiple molecules of a soluble antigen or toxin, clumping whole cells together in a process known as agglutination, or causing soluble antigen molecules to clump together and precipitate out of solution. Neutralization, agglutination, and precipitation of antigens all enhance the likelihood that phagocytotic cells will engulf the antigen (or antigen-bearing cell).
An antibody bound to an antigen molecule on the surface of a pathogen can enhance the phagocytosis of the pathogen. It can also fix and activate the complement system, a series of signaling cascades which lead to an enhancement of phagocytosis, a local inflammatory response, and lysis of the pathogen.
Clonal Selection of B Cells: Clonal selection and expansion work much the same way in B cells as in T cells. Only B cells with appropriate antigen specificity are selected for and expanded (Figure 13). Eventually, the plasma cells secrete antibodies with antigenic specificity identical to those that were on the surfaces of the selected B cells. Notice in the figure that both plasma cells and memory B cells are generated simultaneously.
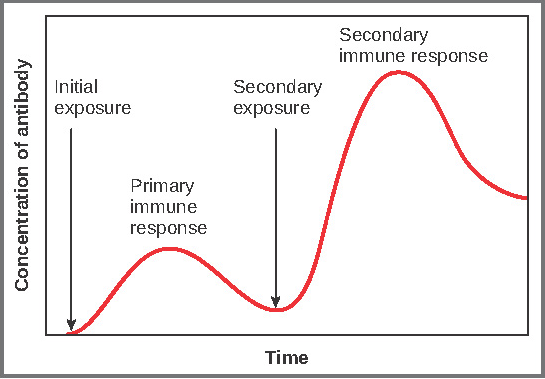
Primary versus Secondary B Cell Responses: Primary and secondary responses as they relate to T cells were discussed earlier. This section will look at these responses with B cells and antibody production. Because antibodies are easily measured in blood samples, their concentrations are easy to follow and graph (Figure 14). As you will see from the figure, the primary response to an antigen (representing a pathogen) is delayed by several days. This is the time it takes for the B cell clones to expand and differentiate into plasma cells. The level of antibody produced is low, but it is sufficient for immune protection. The second time a person encounters the same antigen, there is no time delay, and the amount of antibody made is much higher. Thus, the secondary antibody response overwhelms the pathogens quickly and, in most situations, no symptoms are felt. When a different antigen is used, another primary response is made with its low antibody levels and time delay.
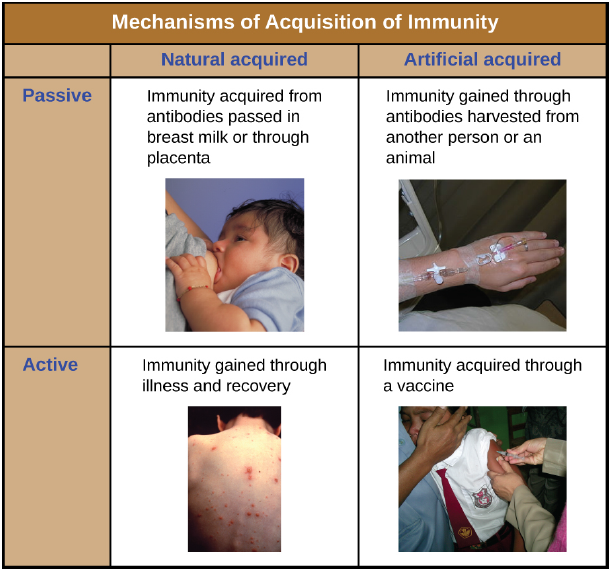
Active versus Passive Immunity: Immunity to pathogens, and the ability to control pathogen growth so that damage to the tissues of the body is limited, can be acquired by (1) the active development of an immune response in the infected individual or (2) the passive transfer of immune components from an immune individual to a nonimmune one. Both active and passive immunity have examples in the natural world and as part of medicine.
Active immunity is the resistance to pathogens acquired during an adaptive immune response within an individual (Figure 15). Naturally acquired active immunity, the response to a pathogen, is the focus of this section. Artificially acquired active immunity involves the use of vaccines. A vaccine is a killed or weakened (attenuated) pathogen or its components that, when administered to a healthy individual, leads to the development of immunological memory (via a weakened primary immune response) without causing much in the way of symptoms. Killed vaccines or inactivated vaccines consist of pathogens that have been killed and so are no longer viable, but still retain antigens that can be recognized and used to mount an immune response. Live attenuated vaccines are generally used when the pathogen involved does not trigger an immune response when introduced in a killed or inactivated state. The virus that causes measles, for example, when introduced in an inactivated form does not confer lasting immunity to measles. However, this same virus can be left viable but modified to render it incapable of producing the symptoms of measles, while still triggering an appropriate immune response. The pathogens contained in a live attenuated vaccine remain viable, but have been rendered harmless or less virulent. Toxoid vaccines include a toxin molecule that has been modified to be harmless but still elicits an immune response against the toxin. The tetanus vaccine for example contains a modified version of the toxin tetanospasmin that is normally released by the bacterium Clostridium tetani. This vaccine triggers the production of anti-tetanospasmin antibodies that confer immunity to the live bacterium’s harmful effects.
A person can also acquire protection from specific pathogens through the administration of pre-formed antibodies, known as passive immunity. Naturally acquired passive immunity is represented by the transfer of antibodies in breast milk or through the placenta that give new born babies protection against some pathogens as they are developing their own immune response. In medicine, artificially acquired passive immunity usually involves injections of immunoglobulins taken from animals previously exposed to a specific pathogen. This treatment is a fast-acting method of temporarily protecting an individual who was possibly exposed to a pathogen. The downside to this treatment is the lack of the development of immunological memory. Once the antibodies are transferred, they are effective for only a limited time before they degrade, so multiple injections may be necessary. An example is the treatment of suspected rabies with postexposure prophylaxis that includes immunoglobulin injections.
From the above, it is readily apparent that with the use of vaccines, one can avoid the damage from disease that results from the first exposure to the pathogen, yet reap the benefits of protection from immunological memory. The advent of vaccines was one of the major medical advances of the twentieth century and led to the eradication of smallpox and the control of many infectious diseases, including polio, measles, and whooping cough.

Watch this Crash Course video for an overview of the adaptive immune response! Direct link: https://youtu.be/2DFN4IBZ3rI
Diseases Associated with Depressed or Overactive Immune Responses: This section is about how the immune system goes wrong. When it goes haywire, and becomes too weak or too strong, it leads to a state of disease. The factors that maintain immunological homeostasis are complex and incompletely understood.
Immunodeficiencies: As you have seen, the immune system is quite complex. It has many pathways using many cell types and signals. Because it is so complex, there are many ways for it to go wrong, and in the case of immunodeficiencies, become weakened. Inherited immunodeficiencies arise from gene mutations that affect specific components of the immune response. There are also acquired immunodeficiencies that result from causes other than inheritance with potentially devastating effects on the immune system, such as infection with HIV.
Inherited Immunodeficiencies: A list of all inherited immunodeficiencies is well beyond the scope of this book. The list is almost as long as the list of cells, proteins, and signaling molecules of the immune system itself. Some deficiencies, such as those for complement, cause only a higher susceptibility to some Gram-negative bacteria. Others are more severe in their consequences. Certainly, the most serious of the inherited immunodeficiencies is severe combined immunodeficiency disease (SCID). This disease is complex because it is caused by many different genetic defects. What groups them together is the fact that both the B cell and T cell arms of the adaptive immune response are affected. Children with this disease usually die of opportunistic infections within their first year of life unless they receive a bone marrow transplant.
Human Immunodeficiency Virus/AIDS: Although many viruses cause suppression of the immune system, only one wipes it out completely, and that is the previously mentioned HIV. The virus is transmitted through semen, vaginal fluids, and blood. There are sometimes, but not always, flu-like symptoms in the first 1 to 2 weeks after infection. Following this time (with no medical intervention), the levels of CD4+ cells, especially helper T cells, decline steadily, eventually producing an acquired immunodeficiency syndrome (AIDS), until at some point, the immune response is so weak that opportunistic disease and eventually death result. CD4 is the receptor that HIV uses to get inside T cells and reproduce. Given that CD4+ helper T cells play an important role in other in T cell immune responses and antibody responses, it should be no surprise that both types of cellular and humoral immune responses are eventually seriously compromised.
Treatment for the disease consists of drugs that target virally encoded proteins that are necessary for viral replication but are absent from normal human cells. By targeting the virus itself and sparing the cells, this approach has been successful in significantly prolonging the lives of HIV-positive individuals. On the other hand, an HIV vaccine has been 30 years in development and is still years away. Because the virus mutates rapidly to evade the immune system, scientists have been looking for parts of the virus that do not change and thus would be good targets for a vaccine candidate.
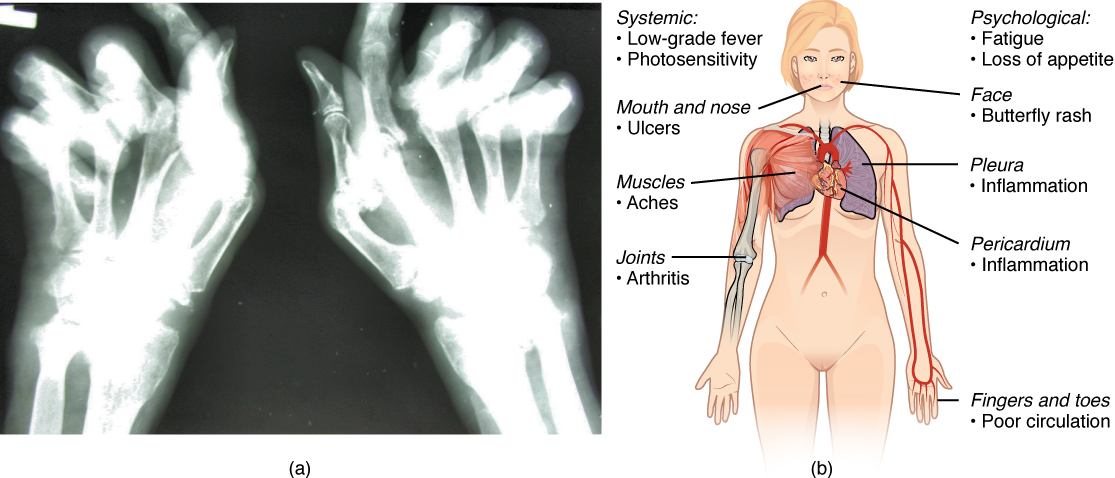
Autoimmune Responses: The worst cases of the immune system over-reacting are autoimmune diseases. Somehow, tolerance breaks down and the immune systems in individuals with these diseases begin to attack their own bodies, causing significant damage. The trigger for these diseases is, more often than not, unknown, and the treatments are usually based on resolving the symptoms using immunosuppressive and anti-inflammatory drugs such as steroids. These diseases can be localized and crippling, as in rheumatoid arthritis, or diffuse in the body with multiple symptoms that differ in different individuals, as is the case with systemic lupus erythematosus (Figure 16).
Part 3: Blood Typing
Blood transfusions in humans were risky procedures until the discovery of the major human blood groups by Karl Landsteiner, an Austrian biologist and physician, in 1900. Until that point, physicians did not understand that death sometimes followed blood transfusions, when the type of donor blood infused into the patient was incompatible with the patient’s own blood. Blood groups are determined by the presence or absence of specific marker molecules on the plasma membranes of erythrocytes. With their discovery, it became possible for the first time to match patient-donor blood types and prevent transfusion reactions and deaths.
Antigens, Antibodies, and Transfusion Reactions: Antigens are substances that the body does not recognize as belonging to the “self” and therefore trigger a defensive response from the leukocytes of the immune system. Here, we will focus on the role of immunity in blood transfusion reactions. Following an infusion of incompatible blood, erythrocytes with foreign antigens appear in the bloodstream and trigger an immune response. Antibodies produced by the plasma cells, attach to the antigens on the plasma membranes of the infused erythrocytes and cause them to adhere to one another.
- As explained before, because the arms of the Y-shaped antibodies attach randomly to more than one non-self erythrocyte surface, they form clumps of erythrocytes (agglutination).
- The clumps of erythrocytes block small blood vessels throughout the body, depriving tissues of oxygen and nutrients.
- As the erythrocyte clumps are degraded, in a process called hemolysis, their hemoglobin is released into the bloodstream. This hemoglobin travels to the kidneys, which are responsible for filtration of the blood. However, the load of hemoglobin released can easily overwhelm the kidney’s capacity to clear it, and the patient can quickly develop kidney failure.
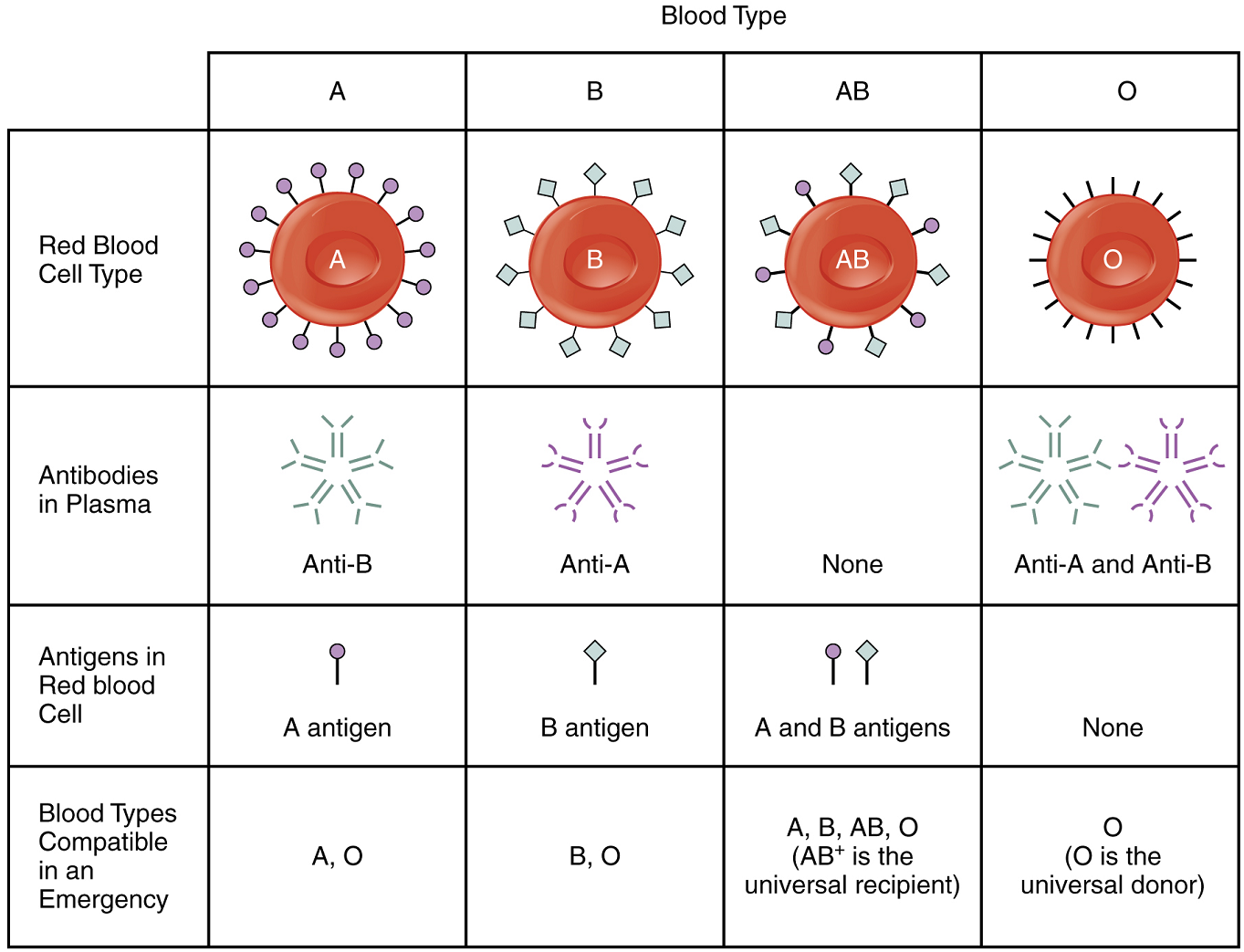
More than 50 antigens have been identified on erythrocyte membranes, but the most significant in terms of their potential harm to patients are classified in two groups: the ABO blood group and the Rh blood group.
The ABO Blood Group: Although the ABO blood group name consists of three letters, ABO blood typing designates the presence or absence of just two antigens, A and B. Both are glycoproteins. People who have A antigens on their erythrocyte membrane surfaces are designated blood type A, and those whose erythrocytes have B antigens are blood type B. People can also have both A and B antigens on their erythrocytes, in which case they are blood type AB. People with neither A nor B antigens are designated blood type O. ABO blood types are genetically determined.
Normally the body must be exposed to a foreign antigen before an antibody can be produced. This is not the case for the ABO blood group. Individuals with type A blood—without any prior exposure to incompatible blood—have pre-formed antibodies to the B antigen circulating in their blood plasma. These antibodies, referred to as anti-B antibodies, will cause agglutination and hemolysis if they ever encounter erythrocytes with B antigens. Similarly, an individual with type B blood has pre-formed anti-A antibodies. Individuals with type AB blood, which has both antigens, do not have pre-formed antibodies to either of these. People with type O blood lack antigens A and B on their erythrocytes, but both pre-formed anti-A and anti-B antibodies circulate in their blood plasma.
Rh Blood Groups: The Rh blood group is classified according to the presence or absence of a second erythrocyte antigen identified as Rh. (It was first discovered in a type of primate known as a rhesus macaque, which is often used in research, because its blood is similar to that of humans.) Although dozens of Rh antigens have been identified, only one, designated D, is clinically important. Those who have the Rh D antigen present on their erythrocytes—about 85 percent of Americans—are described as Rh positive (Rh+) and those who lack it are Rh negative (Rh−). Note that the Rh group is distinct from the ABO group, so any individual, no matter their ABO blood type, may have or lack this Rh antigen. When identifying a patient’s blood type, the Rh group is designated by adding the word positive or negative to the ABO type. For example, A positive (A+) means ABO group A blood with the Rh antigen present, and AB negative (AB−) means ABO group AB blood without the Rh antigen.
In contrast to the ABO group antibodies, which are preformed, antibodies to the Rh antigen are produced only in Rh− individuals after exposure to the antigen. This process, called sensitization, occurs following a transfusion with Rh-incompatible blood or, more commonly, with the birth of an Rh+ baby to an Rh− mother. Problems are rare in a first pregnancy, since the baby’s Rh+ cells rarely cross the placenta (the organ of gas and nutrient exchange between the baby and the mother). However, during or immediately after birth, the Rh− mother can be exposed to the baby’s Rh+ cells (Figure 17). Research has shown that this occurs in about 13−14 percent of such pregnancies. After exposure, the mother’s immune system begins to generate anti-Rh antibodies. If the mother should then conceive another Rh+ baby, the Rh antibodies she has produced can cross the placenta into the fetal bloodstream and destroy the fetal RBCs. This condition, known as hemolytic disease of the newborn (HDN) or erythroblastosis fetalis, may cause anemia in mild cases, but the agglutination and hemolysis can be so severe that without treatment the fetus may die in the womb or shortly after birth.
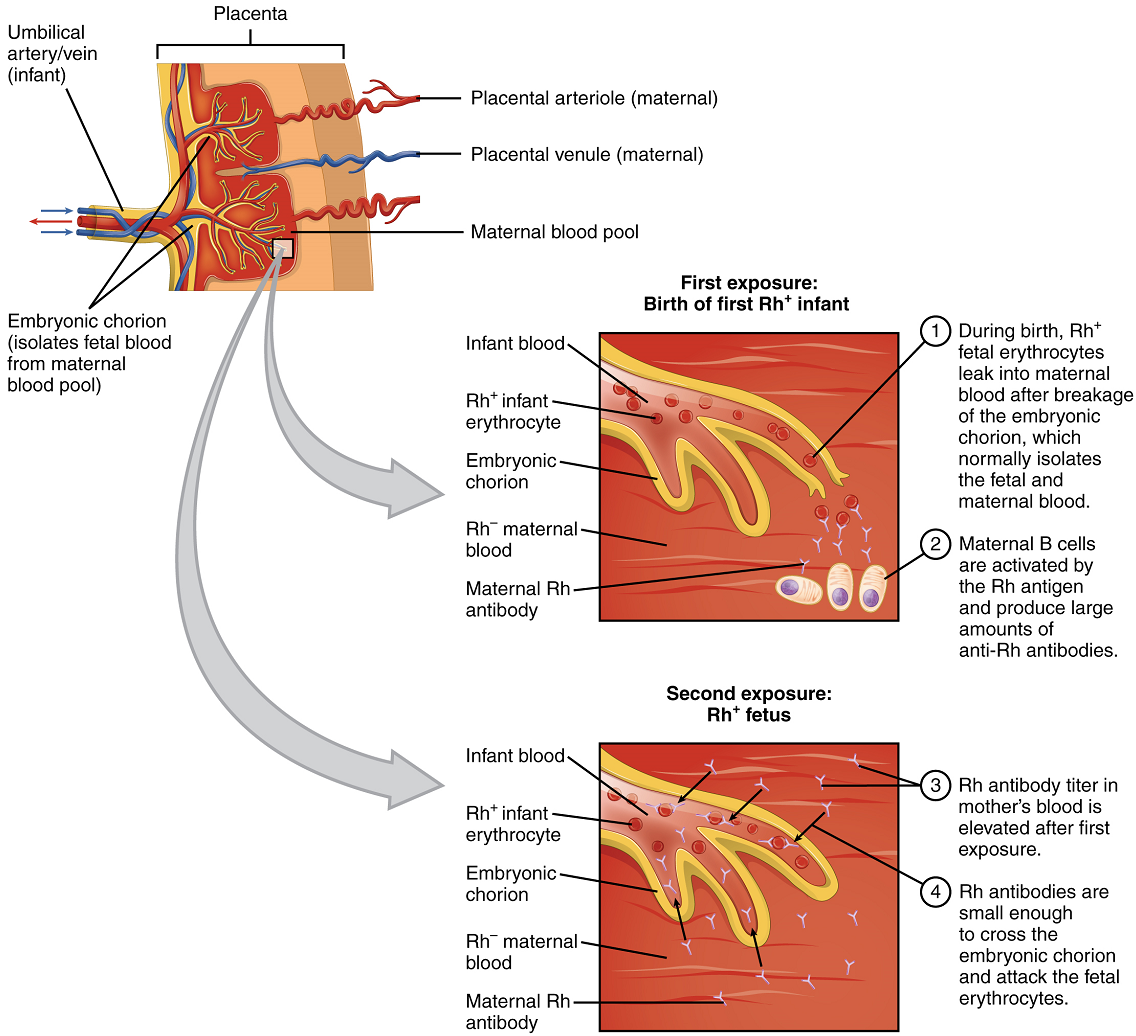
A drug known as RhoGAM, short for Rh immune globulin, can temporarily prevent the development of Rh antibodies in the Rh− mother, thereby averting this potentially serious disease for the fetus. RhoGAM antibodies destroy any fetal Rh+ erythrocytes that may cross the placental barrier. RhoGAM is normally administered to Rh− mothers during weeks 26−28 of pregnancy and within 72 hours following birth. It has proven remarkably effective in decreasing the incidence of HDN. Earlier we noted that the incidence of HDN in an Rh+ subsequent pregnancy to an Rh− mother is about 13–14 percent without preventive treatment. Since the introduction of RhoGAM in 1968, the incidence has dropped to about 0.1 percent in the United States.
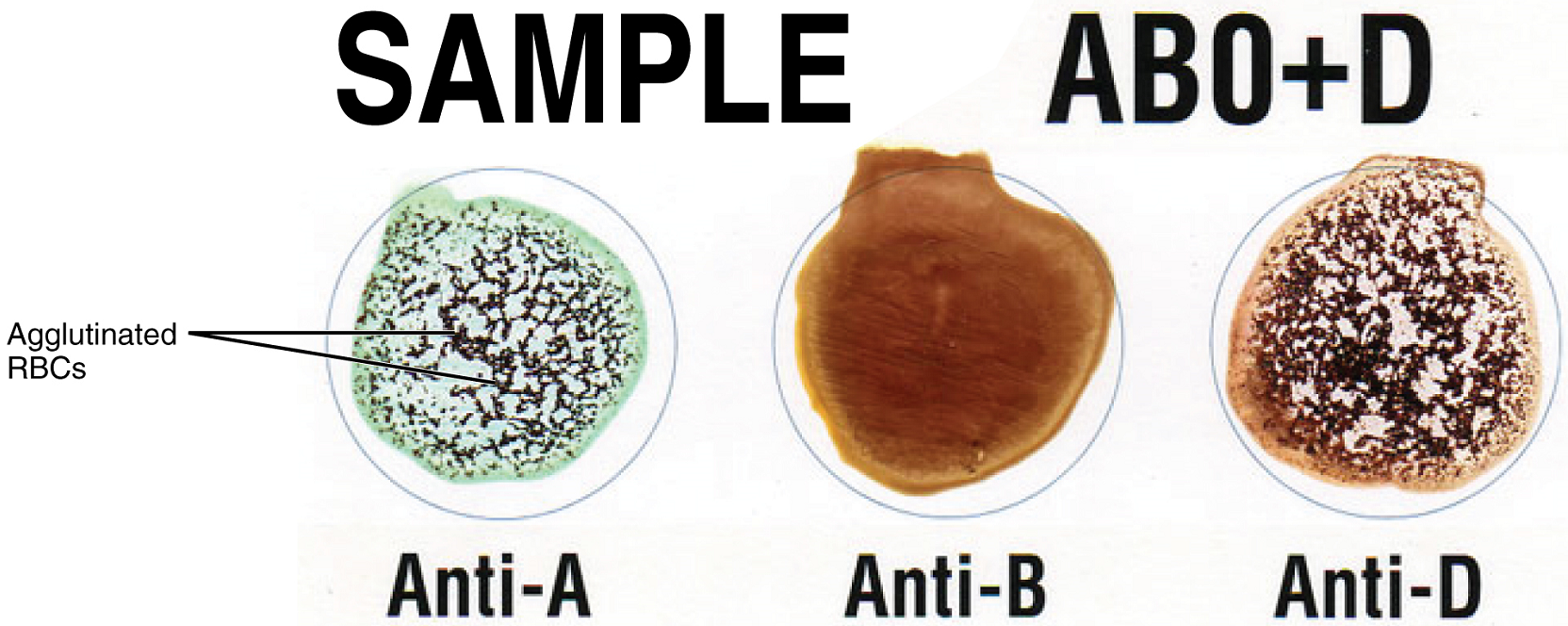
Determining ABO Blood Types: Clinicians are able to determine a patient’s blood type quickly and easily using commercially prepared antibodies. An unknown blood sample is allocated into separate wells. Into one well a small amount of anti-A antibody is added, and to another a small amount of anti-B antibody. If the antigen is present, the antibodies will cause visible agglutination of the cells (Figure 18). The blood should also be tested with Rh antibodies.
ABO Transfusion Protocols: To avoid transfusion reactions, it is best to transfuse only matching blood types; that is, a type B+ recipient should ideally receive blood only from a type B+ donor and so on. That said, in emergency situations, when acute hemorrhage threatens the patient’s life, there may not be time for cross matching to identify blood type. In these cases, blood from a universal donor—an individual with type O− blood—may be transfused. Recall that type O erythrocytes do not display A or B antigens. Thus, anti-A or anti-B antibodies that might be circulating in the patient’s blood plasma will not encounter any erythrocyte surface antigens on the donated blood and therefore will not be provoked into a response.
A patient with blood type AB+ is known as the universal recipient. This patient can theoretically receive any type of blood, because the patient’s own blood—having both A and B antigens on the erythrocyte surface—does not produce anti-A or anti-B antibodies. In addition, an Rh+ patient can receive both Rh+ and Rh− blood. Figure 19 summarizes the blood types and compatibilities.