1103 Chapter 20. The Cardiovascular System: Blood Vessels and Circulation
20.3 Capillary Exchange
Learning Objectives
By the end of this section, you will be able to:
The primary purpose of the cardiovascular system is to circulate gases, nutrients, wastes, and other substances to and from the cells of the body. Small molecules, such as gases, lipids, and lipid-soluble molecules, can diffuse directly through the membranes of the endothelial cells of the capillary wall. Glucose, amino acids, and ions—including sodium, potassium, calcium, and chloride—use transporters to move through specific channels in the membrane by facilitated diffusion. Glucose, ions, and larger molecules may also leave the blood through intercellular clefts. Larger molecules can pass through the pores of fenestrated capillaries, and even large plasma proteins can pass through the great gaps in the sinusoids. Some large proteins in blood plasma can move into and out of the endothelial cells packaged within vesicles by endocytosis and exocytosis. Water moves by osmosis.
Bulk Flow
The mass movement of fluids into and out of capillary beds requires a transport mechanism far more efficient than mere diffusion. This movement, often referred to as bulk flow, involves two pressure-driven mechanisms: Volumes of fluid move from an area of higher pressure in a capillary bed to an area of lower pressure in the tissues via filtration. In contrast, the movement of fluid from an area of higher pressure in the tissues into an area of lower pressure in the capillaries is reabsorption. Two types of pressure interact to drive each of these movements: hydrostatic pressure and osmotic pressure.
Hydrostatic Pressure
The primary force driving fluid transport between the capillaries and tissues is hydrostatic pressure, which can be defined as the pressure of any fluid enclosed in a space. Blood hydrostatic pressure is the force exerted by the blood confined within blood vessels or heart chambers. Even more specifically, the pressure exerted by blood against the wall of a capillary is called capillary hydrostatic pressure (CHP), and is the same as capillary blood pressure. CHP is the force that drives fluid out of capillaries and into the tissues.
As fluid exits a capillary and moves into tissues, the hydrostatic pressure in the interstitial fluid correspondingly rises. This opposing hydrostatic pressure is called the interstitial fluid hydrostatic pressure (IFHP). Generally, the CHP originating from the arterial pathways is considerably higher than the IFHP, because lymphatic vessels are continually absorbing excess fluid from the tissues. Thus, fluid generally moves out of the capillary and into the interstitial fluid. This process is called filtration.
Osmotic Pressure
The net pressure that drives reabsorption—the movement of fluid from the interstitial fluid back into the capillaries—is called osmotic pressure (sometimes referred to as oncotic pressure). Whereas hydrostatic pressure forces fluid out of the capillary, osmotic pressure draws fluid back in. Osmotic pressure is determined by osmotic concentration gradients, that is, the difference in the solute-to-water concentrations in the blood and tissue fluid. A region higher in solute concentration (and lower in water concentration) draws water across a semipermeable membrane from a region higher in water concentration (and lower in solute concentration).
As we discuss osmotic pressure in blood and tissue fluid, it is important to recognize that the formed elements of blood do not contribute to osmotic concentration gradients. Rather, it is the plasma proteins that play the key role. Solutes also move across the capillary wall according to their concentration gradient, but overall, the concentrations should be similar and not have a significant impact on osmosis. Because of their large size and chemical structure, plasma proteins are not truly solutes, that is, they do not dissolve but are dispersed or suspended in their fluid medium, forming a colloid rather than a solution.
The pressure created by the concentration of colloidal proteins in the blood is called the blood colloidal osmotic pressure (BCOP). Its effect on capillary exchange accounts for the reabsorption of water. The plasma proteins suspended in blood cannot move across the semipermeable capillary cell membrane, and so they remain in the plasma. As a result, blood has a higher colloidal concentration and lower water concentration than tissue fluid. It therefore attracts water. We can also say that the BCOP is higher than the interstitial fluid colloidal osmotic pressure (IFCOP), which is always very low because interstitial fluid contains few proteins. Thus, water is drawn from the tissue fluid back into the capillary, carrying dissolved molecules with it. This difference in colloidal osmotic pressure accounts for reabsorption.
Interaction of Hydrostatic and Osmotic Pressures
The normal unit used to express pressures within the cardiovascular system is millimeters of mercury (mm Hg). When blood leaving an arteriole first enters a capillary bed, the CHP is quite high—about 35 mm Hg. Gradually, this initial CHP declines as the blood moves through the capillary so that by the time the blood has reached the venous end, the CHP has dropped to approximately 18 mm Hg. In comparison, the plasma proteins remain suspended in the blood, so the BCOP remains fairly constant at about 25 mm Hg throughout the length of the capillary and considerably below the osmotic pressure in the interstitial fluid.
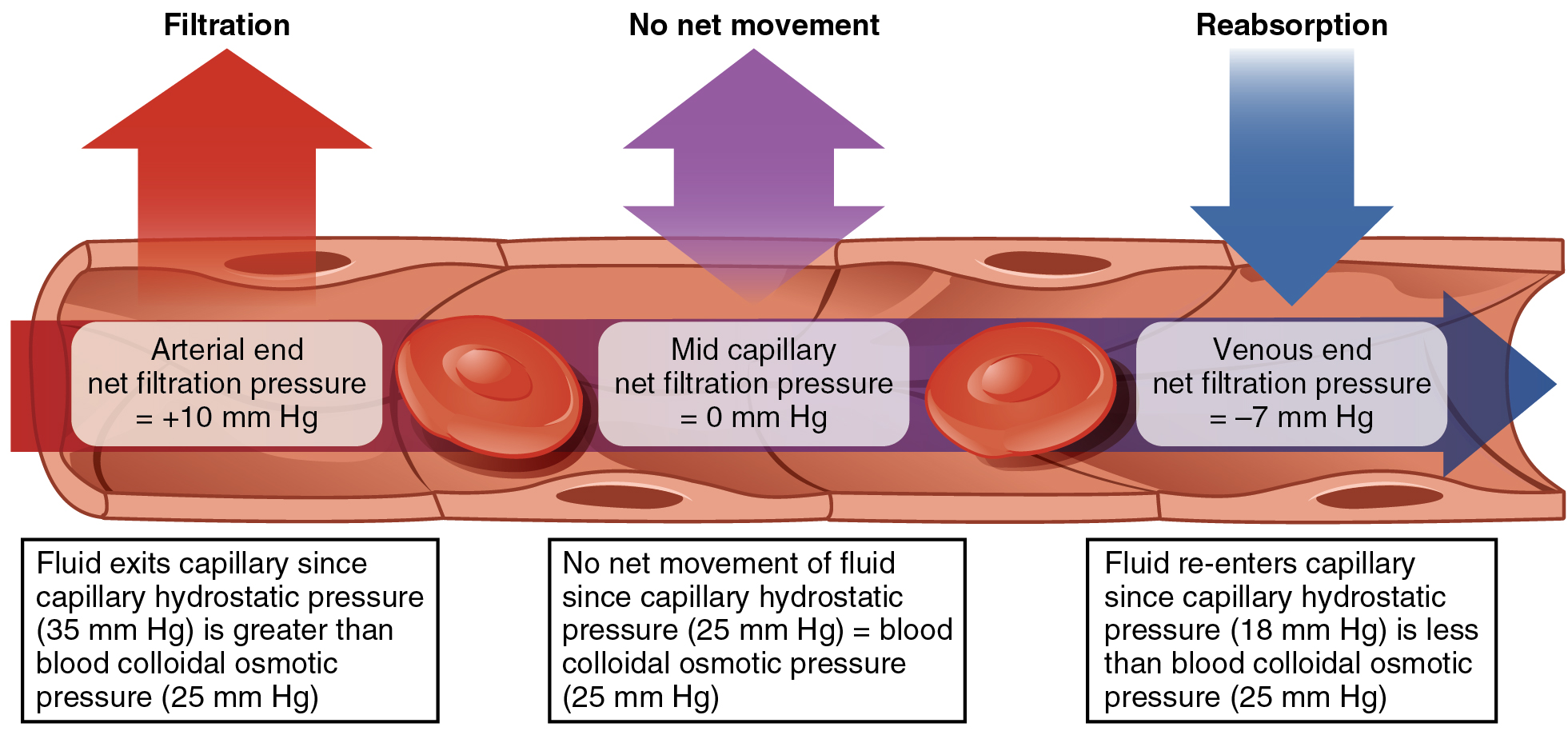
The net filtration pressure (NFP) represents the interaction of the hydrostatic and osmotic pressures, driving fluid out of the capillary. It is equal to the difference between the CHP and the BCOP. Since filtration is, by definition, the movement of fluid out of the capillary, when reabsorption is occurring, the NFP is a negative number.
NFP changes at different points in a capillary bed (Figure 1). Close to the arterial end of the capillary, it is approximately 10 mm Hg, because the CHP of 35 mm Hg minus the BCOP of 25 mm Hg equals 10 mm Hg. Recall that the hydrostatic and osmotic pressures of the interstitial fluid are essentially negligible. Thus, the NFP of 10 mm Hg drives a net movement of fluid out of the capillary at the arterial end. At approximately the middle of the capillary, the CHP is about the same as the BCOP of 25 mm Hg, so the NFP drops to zero. At this point, there is no net change of volume: Fluid moves out of the capillary at the same rate as it moves into the capillary. Near the venous end of the capillary, the CHP has dwindled to about 18 mm Hg due to loss of fluid. Because the BCOP remains steady at 25 mm Hg, water is drawn into the capillary, that is, reabsorption occurs. Another way of expressing this is to say that at the venous end of the capillary, there is an NFP of −7 mm Hg.