1103 Chapter 21. The Lymphatic and Immune System
21.5 The Immune Response against Pathogens
Learning Objectives
By the end of this section, you will be able to:
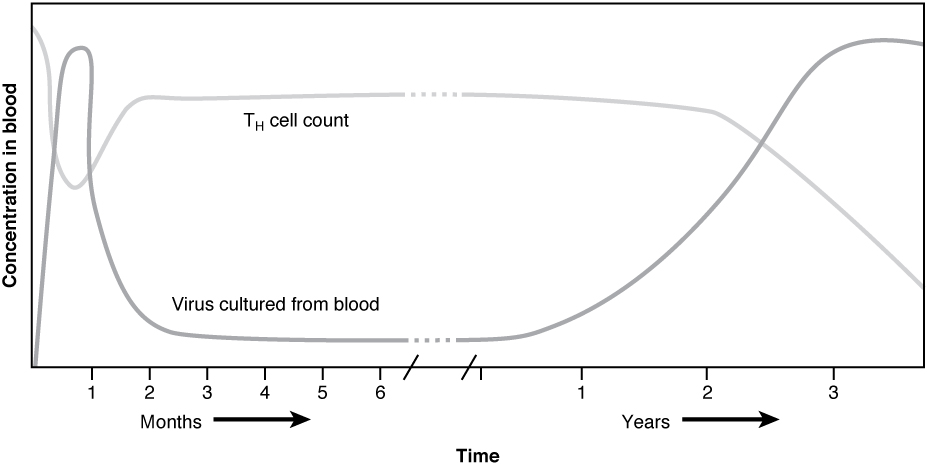
Now that you understand the development of mature, naïve B cells and T cells, and some of their major functions, how do all of these various cells, proteins, and cytokines come together to actually resolve an infection? Ideally, the immune response will rid the body of a pathogen entirely. The adaptive immune response, with its rapid clonal expansion, is well suited to this purpose. Think of a primary infection as a race between the pathogen and the immune system. The pathogen bypasses barrier defenses and starts multiplying in the host’s body. During the first 4 to 5 days, the innate immune response will partially control, but not stop, pathogen growth. As the adaptive immune response gears up, however, it will begin to clear the pathogen from the body, while at the same time becoming stronger and stronger. When following antibody responses in patients with a particular disease such as a virus, this clearance is referred to as seroconversion (sero- = “serum”). Seroconversion is the reciprocal relationship between virus levels in the blood and antibody levels. As the antibody levels rise, the virus levels decline, and this is a sign that the immune response is being at least partially effective (partially, because in many diseases, seroconversion does not necessarily mean a patient is getting well).
An excellent example of this is seroconversion during HIV disease (Figure 1). Notice that antibodies are made early in this disease, and the increase in anti-HIV antibodies correlates with a decrease in detectable virus in the blood. Although these antibodies are an important marker for diagnosing the disease, they are not sufficient to completely clear the virus. Several years later, the vast majority of these individuals, if untreated, will lose their entire adaptive immune response, including the ability to make antibodies, during the final stages of AIDS.
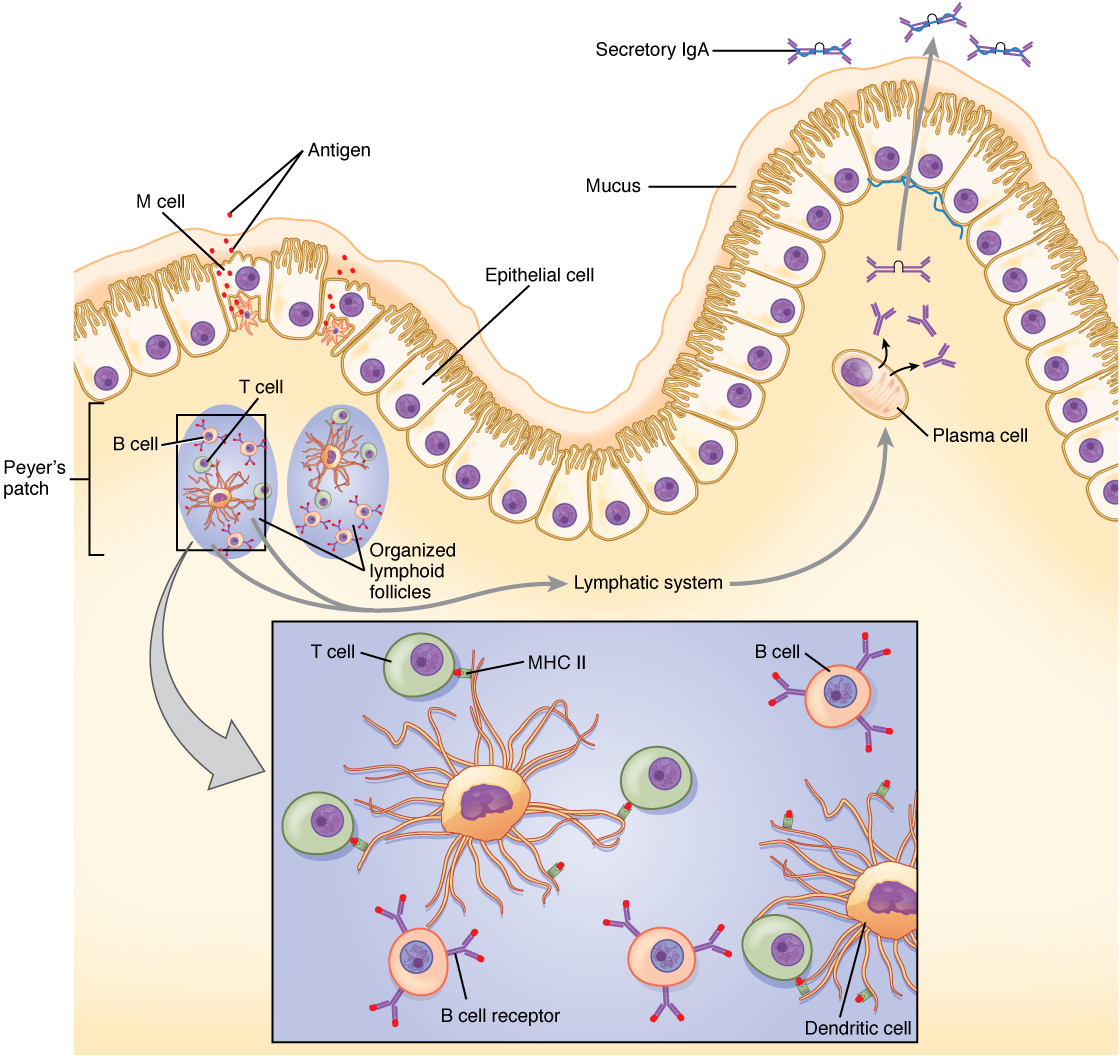
The Mucosal Immune Response
Mucosal tissues are major barriers to the entry of pathogens into the body. The IgA (and sometimes IgM) antibodies in mucus and other secretions can bind to the pathogen, and in the cases of many viruses and bacteria, neutralize them. Neutralization is the process of coating a pathogen with antibodies, making it physically impossible for the pathogen to bind to receptors. Neutralization, which occurs in the blood, lymph, and other body fluids and secretions, protects the body constantly. Neutralizing antibodies are the basis for the disease protection offered by vaccines. Vaccinations for diseases that commonly enter the body via mucous membranes, such as influenza, are usually formulated to enhance IgA production.
Immune responses in some mucosal tissues such as the Peyer’s patches (see Chapter 21.1 Figure 10) in the small intestine take up particulate antigens by specialized cells known as microfold or M cells (Figure 2). These cells allow the body to sample potential pathogens from the intestinal lumen. Dendritic cells then take the antigen to the regional lymph nodes, where an immune response is mounted.
Defenses against Bacteria and Fungi
The body fights bacterial pathogens with a wide variety of immunological mechanisms, essentially trying to find one that is effective. Bacteria such as Mycobacterium leprae, the cause of leprosy, are resistant to lysosomal enzymes and can persist in macrophage organelles or escape into the cytosol. In such situations, infected macrophages receiving cytokine signals from Th1 cells turn on special metabolic pathways. Macrophage oxidative metabolism is hostile to intracellular bacteria, often relying on the production of nitric oxide to kill the bacteria inside the macrophage.
Fungal infections, such as those from Aspergillus, Candida, and Pneumocystis, are largely opportunistic infections that take advantage of suppressed immune responses. Most of the same immune mechanisms effective against bacteria have similar effects on fungi, both of which have characteristic cell wall structures that protect their cells.
Defenses against Parasites
Worm parasites such as helminths are seen as the primary reason why the mucosal immune response, IgE-mediated allergy and asthma, and eosinophils evolved. These parasites were at one time very common in human society. When infecting a human, often via contaminated food, some worms take up residence in the gastrointestinal tract. Eosinophils are attracted to the site by T cell cytokines, which release their granule contents upon their arrival. Mast cell degranulation also occurs, and the fluid leakage caused by the increase in local vascular permeability is thought to have a flushing action on the parasite, expelling its larvae from the body. Furthermore, if IgE labels the parasite, the eosinophils can bind to it by its Fc receptor.
Defenses against Viruses
The primary mechanisms against viruses are NK cells, interferons, and cytotoxic T cells. Antibodies are effective against viruses mostly during protection, where an immune individual can neutralize them based on a previous exposure. Antibodies have no effect on viruses or other intracellular pathogens once they enter the cell, since antibodies are not able to penetrate the plasma membrane of the cell. Many cells respond to viral infections by downregulating their expression of MHC class I molecules. This is to the advantage of the virus, because without class I expression, cytotoxic T cells have no activity. NK cells, however, can recognize virally infected class I-negative cells and destroy them. Thus, NK and cytotoxic T cells have complementary activities against virally infected cells.
Interferons have activity in slowing viral replication and are used in the treatment of certain viral diseases, such as hepatitis B and C, but their ability to eliminate the virus completely is limited. The cytotoxic T cell response, though, is key, as it eventually overwhelms the virus and kills infected cells before the virus can complete its replicative cycle. Clonal expansion and the ability of cytotoxic T cells to kill more than one target cell make these cells especially effective against viruses. In fact, without cytotoxic T cells, it is likely that humans would all die at some point from a viral infection (if no vaccine were available).
Evasion of the Immune System by Pathogens
It is important to keep in mind that although the immune system has evolved to be able to control many pathogens, pathogens themselves have evolved ways to evade the immune response. An example already mentioned is in Mycobactrium tuberculosis, which has evolved a complex cell wall that is resistant to the digestive enzymes of the macrophages that ingest them, and thus persists in the host, causing the chronic disease tuberculosis. This section briefly summarizes other ways in which pathogens can “outwit” immune responses. But keep in mind, although it seems as if pathogens have a will of their own, they do not. All of these evasive “strategies” arose strictly by evolution, driven by selection.
Bacteria sometimes evade immune responses because they exist in multiple strains, such as different groups of Staphylococcus aureus. S. aureus is commonly found in minor skin infections, such as boils, and some healthy people harbor it in their nose. One small group of strains of this bacterium, however, called methicillin-resistant Staphylococcus aureus, has become resistant to multiple antibiotics and is essentially untreatable. Different bacterial strains differ in the antigens on their surfaces. The immune response against one strain (antigen) does not affect the other; thus, the species survives.
Another method of immune evasion is mutation. Because viruses’ surface molecules mutate continuously, viruses like influenza change enough each year that the flu vaccine for one year may not protect against the flu common to the next. New vaccine formulations must be derived for each flu season.
Genetic recombination—the combining of gene segments from two different pathogens—is an efficient form of immune evasion. For example, the influenza virus contains gene segments that can recombine when two different viruses infect the same cell. Recombination between human and pig influenza viruses led to the 2010 H1N1 swine flu outbreak.
Pathogens can produce immunosuppressive molecules that impair immune function, and there are several different types. Viruses are especially good at evading the immune response in this way, and many types of viruses have been shown to suppress the host immune response in ways much more subtle than the wholesale destruction caused by HIV.
