1103 Chapter 7. Axial Skeleton
7.5 Embryonic Development of the Axial Skeleton
Learning Objectives
By the end of this section, you will be able to:
- Describe the major differences between the axial skeleton of a baby and that of an adult
The axial skeleton begins to form during early embryonic development. However, growth, remodeling, and ossification (bone formation) continue for several decades after birth before the adult skeleton is fully formed. Knowledge of the developmental processes that give rise to the skeleton is important for understanding the abnormalities that may arise in skeletal structures.
Development of the Skull
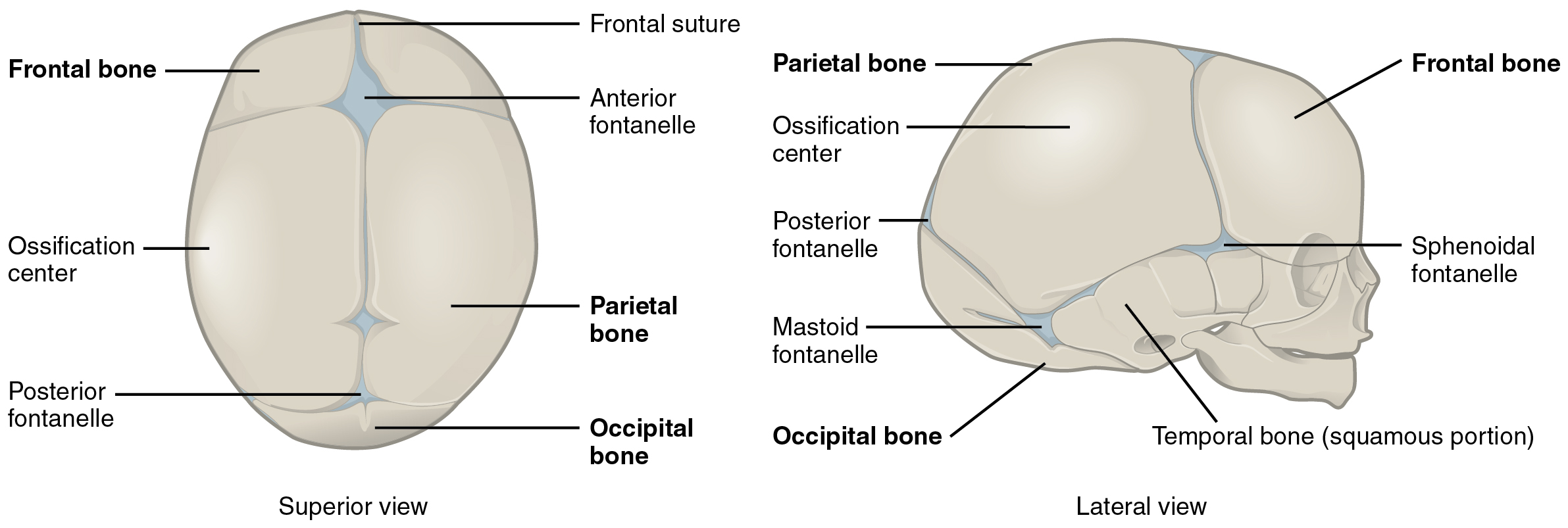
The bones of the skull arise from mesenchyme during embryonic development in two different ways. The first mechanism produces the bones that form the top and sides of the brain case. This involves the local accumulation of mesenchymal cells at the site of the future bone. These cells then differentiate directly into bone producing cells, which form the skull bones through the process of intramembranous ossification. As the brain case bones grow in the fetal skull, they remain separated from each other by large areas of dense connective tissue, each of which is called a fontanelle (Figure 1). The fontanelles are the soft spots on an infant’s head. They are important during birth because these areas allow the skull to change shape as it squeezes through the birth canal. After birth, the fontanelles allow for continued growth and expansion of the skull as the brain enlarges. The largest fontanelle is located on the anterior head, at the junction of the frontal and parietal bones. The fontanelles decrease in size and disappear by age 2. However, the skull bones remained separated from each other at the sutures, which contain dense fibrous connective tissue that unites the adjacent bones. The connective tissue of the sutures allows for continued growth of the skull bones as the brain enlarges during childhood growth. This structure also means that, although the size of the cranium increases from birth to adulthood, proportionately it does so less than other parts of the skeleton; the relative size of the cranium in proportion to the rest of the body therefore decreases with age from birth to adulthood.
The second mechanism for bone development in the skull produces the facial bones and floor of the brain case. This also begins with the localized accumulation of mesenchymal cells. However, these cells differentiate into cartilage cells, which produce a hyaline cartilage model of the future bone. As this cartilage model grows, it is gradually converted into bone through the process of endochondral ossification. This is a slow process and the cartilage is not completely converted to bone until the skull achieves its full adult size.
At birth, the brain case and orbits of the skull are disproportionally large compared to the bones of the jaws and lower face. This reflects the relative underdevelopment of the maxilla and mandible, which lack teeth, and the small sizes of the paranasal sinuses and nasal cavity. During early childhood, the mastoid process enlarges, the two halves of the mandible and frontal bone fuse together to form single bones, and the paranasal sinuses enlarge. The jaws also expand as the teeth begin to appear. These changes all contribute to the rapid growth and enlargement of the face during childhood.
Development of the Vertebral Column and Thoracic cage
Development of the vertebrae begins with the accumulation of mesenchyme cells from each sclerotome around the notochord. These cells differentiate into a hyaline cartilage model for each vertebra, which then grow and eventually ossify into bone through the process of endochondral ossification. As the developing vertebrae grow, the notochord largely disappears. However, small areas of notochord tissue persist between the adjacent vertebrae and this contributes to the formation of each intervertebral disc.

The ribs and sternum also develop from mesenchyme. The ribs initially develop as part of the cartilage model for each vertebra, but in the thorax region, the rib portion separates from the vertebra by the eighth week. The cartilage model of the rib then ossifies, except for the anterior portion, which remains as the costal cartilage. The sternum initially forms as paired hyaline cartilage models on either side of the anterior midline, beginning during the fifth week of development. The cartilage models of the ribs become attached to the lateral sides of the developing sternum. Eventually, the two halves of the cartilaginous sternum fuse together along the midline and then ossify into bone. The manubrium and body of the sternum are converted into bone first, with the xiphoid process remaining as cartilage until late in life.
