Chapter 2 Innate and Adaptive Immunity: From Cell Defense to Tissue Repair
Section 8: Adaptive (Specific) Immunity – T Cells and B Cells
Zoë Soon
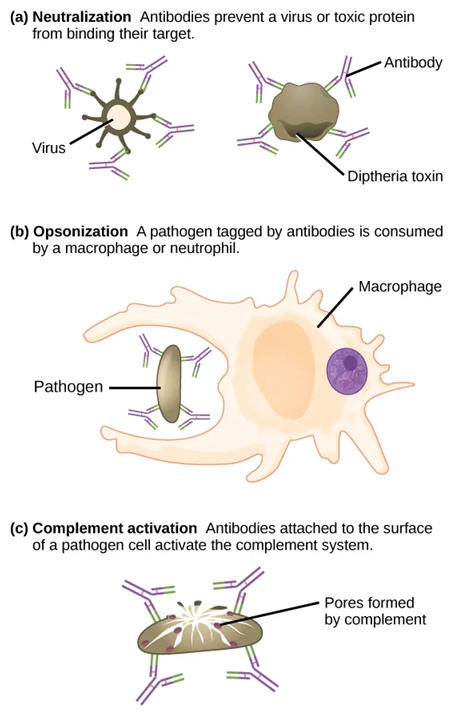
While innate immunity provides immediate, broad protection, adaptive immunity targets specific pathogens and builds long-lasting memory through the production of both Memory T and Memory B cells. Adaptive immunity takes longer to become established than innate immunity as a series of events is required to fully activate the T and B cells responsible for adaptive immunity. The T cells once activated provide cell-mediated immunity in which pathogens are killed directly. B cells provide humoral immunity which involves the production of antibodies that have 3 main roles: (1) neutralizing toxins and pathogens by binding them and preventing contact with healthy cells, (2) opsonization and enhancing the phagocytosis of pathogens, and (3) activating the complement system (and therefore also the inflammatory response).
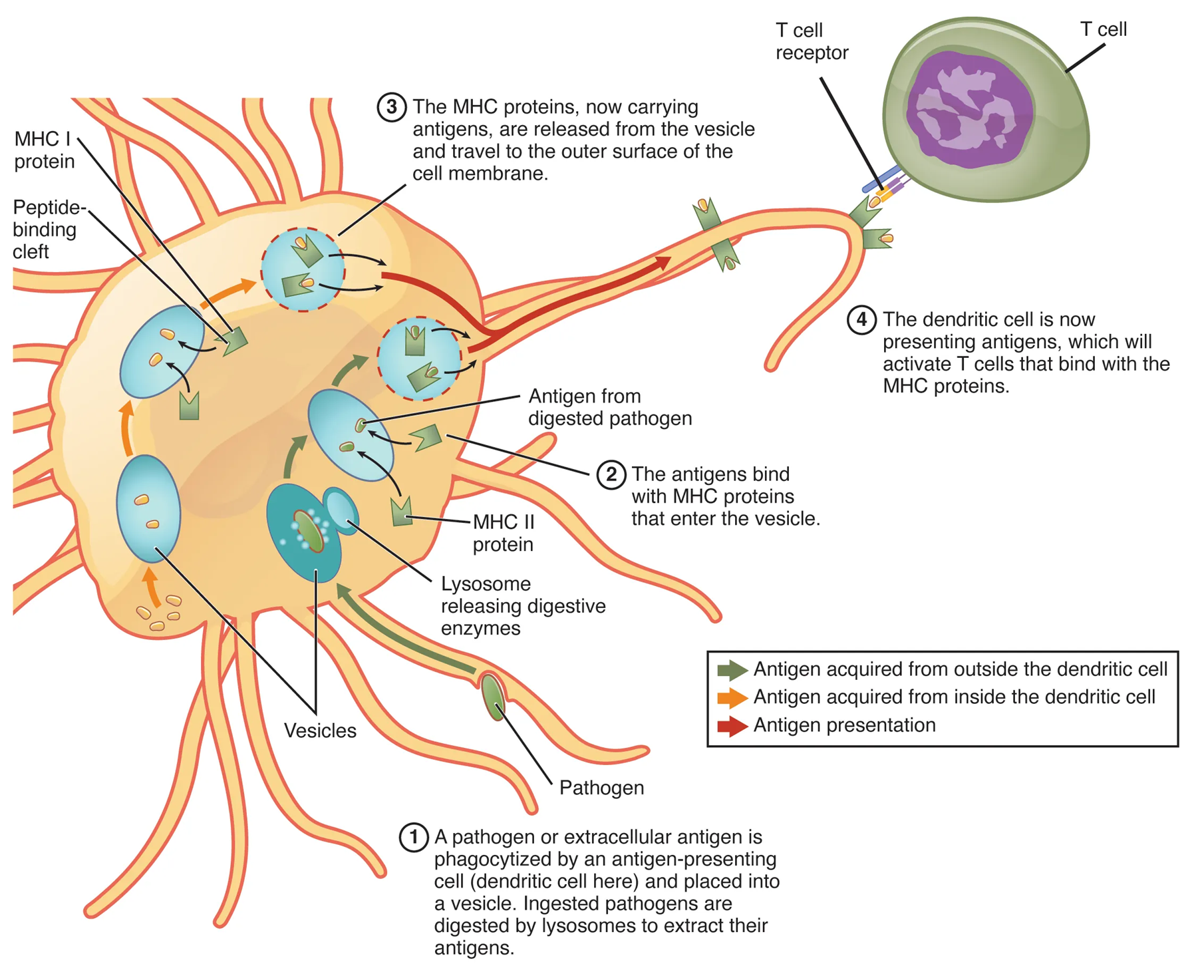
Antigen-Presenting Cells
Antigen-presenting cells (APC) include macrophages, dendritic cells, and B cells, all of which are able to phagocytose and digest pathogens, producing many pathogen fragments (antigens) that can then be displayed on their cell surfaces in order to stimulate the adaptive immune response by both B and T cells.

CD4+ Helper T Cells: Orchestrating the Adaptive Immune Response
Once the APC has engulfed a pathogen, it matures and migrates to a lymph node to activate CD4+ Helper T cells (Th cells) as well as CD8+ Cytotoxic cells (Tc cells). Activated Th cells are responsible for orchestrating the adaptive immune response and play crucial roles in activating both B cells and Tc cells.
To activate Th cells, three signals are required:
| Signal 1:
Non-Self Antigen Presentation |
Th cells with a complementary TCR and CD4 co-receptors bind to the displayed pathogen antigen – MHC-II complexes on the surface of the APC.
The match of TCR to this abnormal antigen and simultaneous confirmation by CD4 binding to the MHC-II portion and stabilization of the bound complex is considered the 1st signal. |
| Signal 2:
Ligand-Receptor Interaction |
Th cells also have CD28 receptors that need to bind with ligands (CD80 and CD86) on the surface of the APC. This “second handshake” is considered the 2nd signal. |
| Signal 3:
Cytokine stimulation |
The APC then releases cytokines to stimulate Th cells to become fully active and proliferate, producing active daughter Th cells (through clonal expansion) that circulate and activate Tc and B cells.
Note: While the APC is activating the Th cell, the Th cell signals feedback to the APC known as licensing, which causes the APC to become even more active and upregulate ligands (CD80 and CD86) that it uses to bind and activate Tc cells. |
*The three signals act as safety checks – if a T cell receives Signal 1, but not Signal 2, it will either become anergic (unresponsive) or undergoes apoptosis to prevent accidental autoimmunity.
Th cells that are activated and proliferate produce the following daughter cell types:
- Effector Th cells: Short-lived cells that leave the lymph node and circulate and activate macrophages, Tc and B cells.
- Memory Th cells: Cells that are able to undergo cell division and retain antigen memory by creating and maintaining a stable pool of Memory Th cells for 10+ years; enabling a faster secondary response upon re-exposure to the same pathogens.
Effector Th cells are successfully activated, they have 3 major roles:
(1) enhance the activation of Tc cells by releasing cytokines.
(2) activate B cells – by binding to sensitized B cell that are displaying pathogenic antigen and releasing cytokines that stimulate the B cell to proliferate producing two types of daughter cells: plasma cells that become antibody factories and memory B cells.
(3) release cytokines to activate macrophages, enhancing their ability to kill pathogens.
CD8+ Cytotoxic T Cells: Search and Destroy
Typically in host cells, as internal proteins are recycled by proteasomes, their fragments are displayed as self-antigens on their MHC-I molecules. This allows for Natural Killer (NK) cells to perform immunological surveillance, an innate immune process by which cells that are not expressing self-antigens are eliminated.
However, when a host cell is infected or abnormal (e.g., cancerous), it displays pathogen/tumor (non-self) antigens via MHC Class I on its surface.
Additionally macrophages and dendritic cells will engulf these pathogens (or cancer cells), becoming APCs and displaying non-self antigens via MHC Class II on their surfaces. If an APC is itself infected, it will present the pathogen antigens using MHC Class I molecules. In special scenarios, APCs will use both MHC I and MHC I to display pathogen/tumor antigens.
Similarly to CD4+ Helper T cells, in order to respond to an infection, CD8+ Cytotoxic T cells require 3 signals in order to become fully activated:
| Signal 1:
Non-Self Antigen Presentation |
Tc cells with a complementary TCR and CD8 co-receptors bind to the displayed pathogen/tumor antigen – MHC-I complexes on the surface of either an infected host cell or an infected APC.
The match of TCR to this non-self antigen and simultaneous confirmation by CD8 binding to the MHC-I portion and stabilization of the bound complex is considered the 1st signal. |
| Signal 2:
Ligand-Receptor Interaction
Cross-bridging
|
The CD28 receptors on Tc cells also need to bind with ligands (CD80 and CD86) that appear on the surface of the activated APC.
Note that this process is greatly enhanced when APCs displays high levels of these ligands, when licensed (further activated by Th cells (as mentioned above). Licensing is an added safety check in the activation of Tc cells, preventing auto-immunity. Cross-bridging can occur between all 3 cells, where an APC can bind both a Th cell and a Tc cell at the same time. In this case, APC displays non-self antigens on MHC-1 molecules that bind to the TCR and CD8 of the Tc cell. At the same time, the APC has displayed non-self antigens on MHC-II that bind to the TCR and CD4 of the Th cell. Both Th and Tc cells must recognize the exact same foreign antigen. This close proximity facilitates cytokine release and stimulation. |
| Signal 3:
Cytokine stimulation |
Cytokines are released from both activated APC and Th cells to stimulate Tc cells to become active, proliferate through clonal expansion and perform seek and destroy missions.
Seeking out and destroying the infected/abnormal cells involves releasing either toxic granules (perforin/granzymes to lyse target cells or inducing target cell-death receptors to trigger apoptosis. |
Tc cells that are activated and proliferate produce two daughter cell types:
- Effector Tc cells: Short-lived cells that circulate and destroy infected/cancerous cells by releasing perforin, granzymes, and other toxins – perforins create membrane portals; cells swell and burst (cytolysis) while granzymes induce apoptosis. This is the ‘Seek and Destroy’ Mission.
- Memory Tc cells: Cells that are able to undergo cell division and retain antigen memory by creating and maintaining a stable pool of Memory Tc cells for 10+ years; enabling a faster secondary response upon re-exposure to the same pathogens.
B Cell Activation and Antibody Production
B cells arise from hemocytoblasts in the bone marrow and reside in secondary lymphatic tissues as self-tolerant naïve B cells. To become active, a B cell must be first sensitized by non-self antigens and then stimulated by active Th cells.
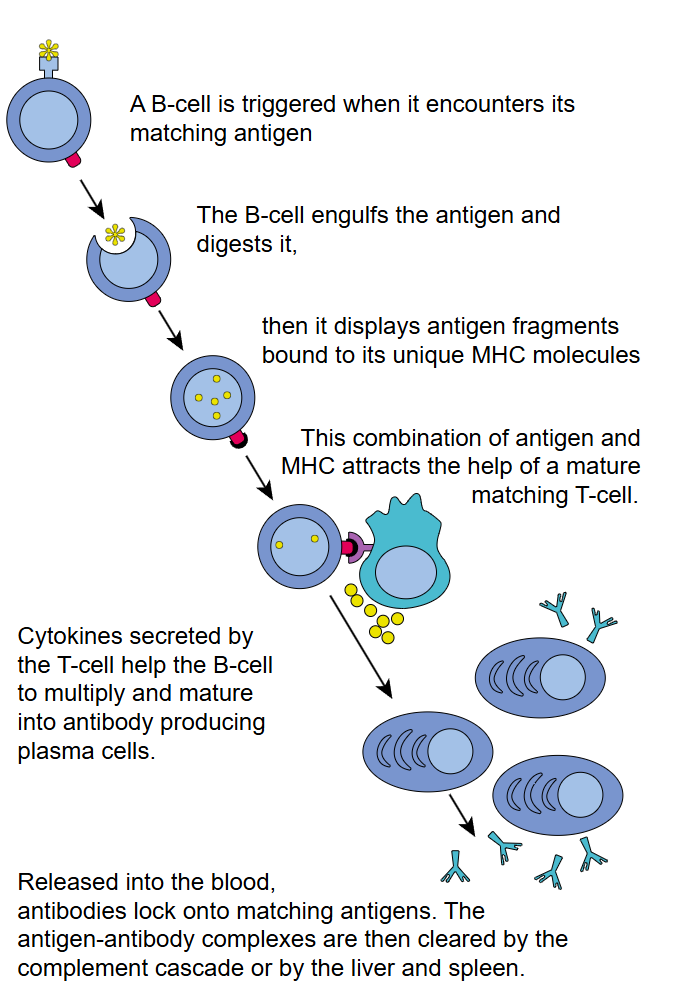
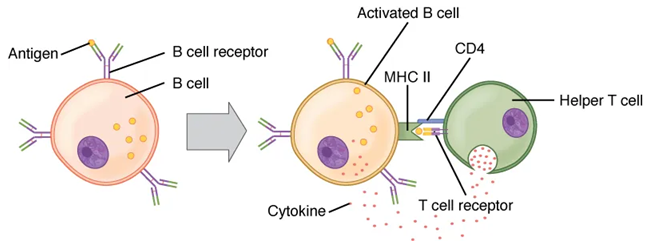
B cells that are activated and proliferate produce two daughter cell types:
- Plasma cells: Secrete antibodies specific to the pathogen antigen. Antibodies circulate, inactivate pathogens, act as opsonins, neutralize toxins, prevent viral entry, and agglutinate pathogens.
- Memory B cells: Provide long-term immunity – mount a faster, stronger antibody response upon re-exposure.
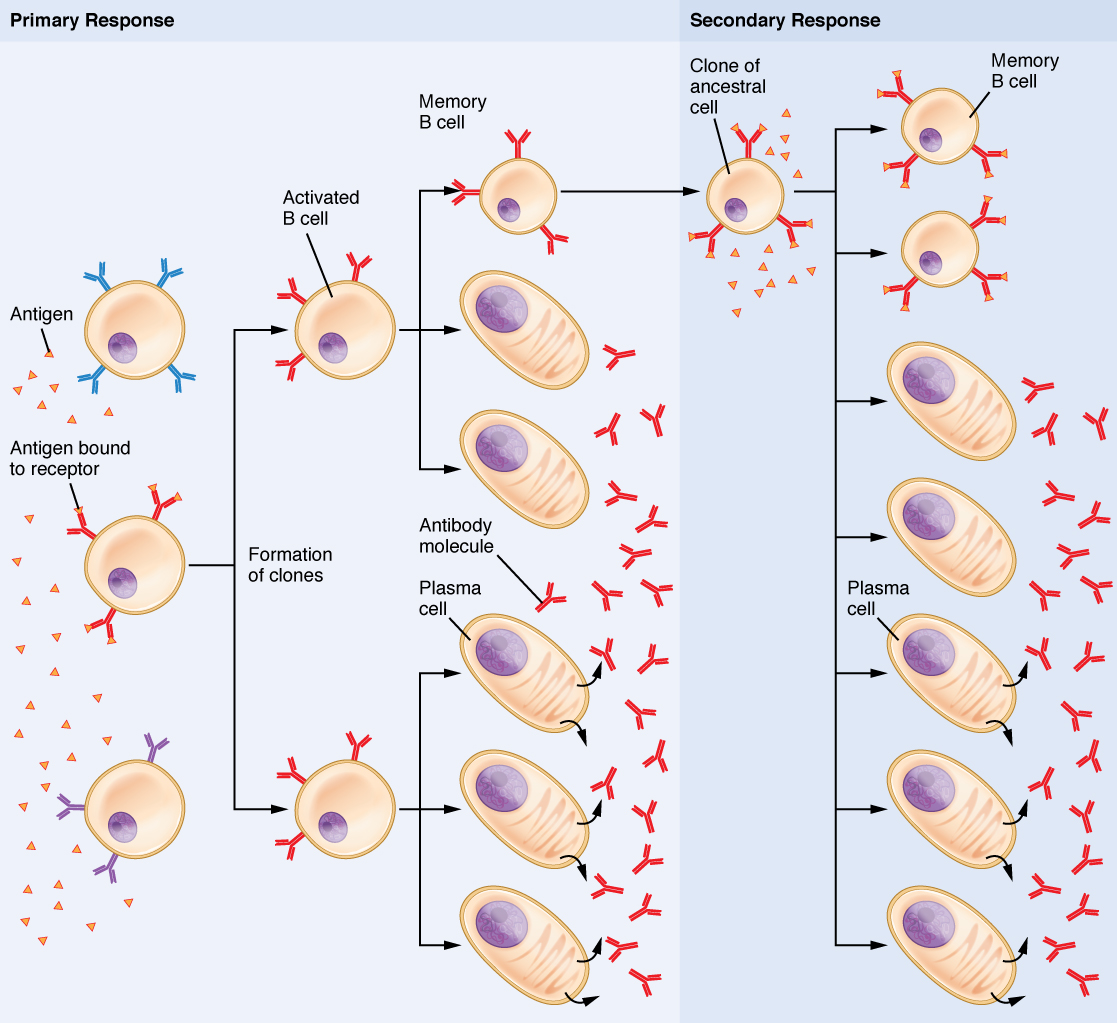
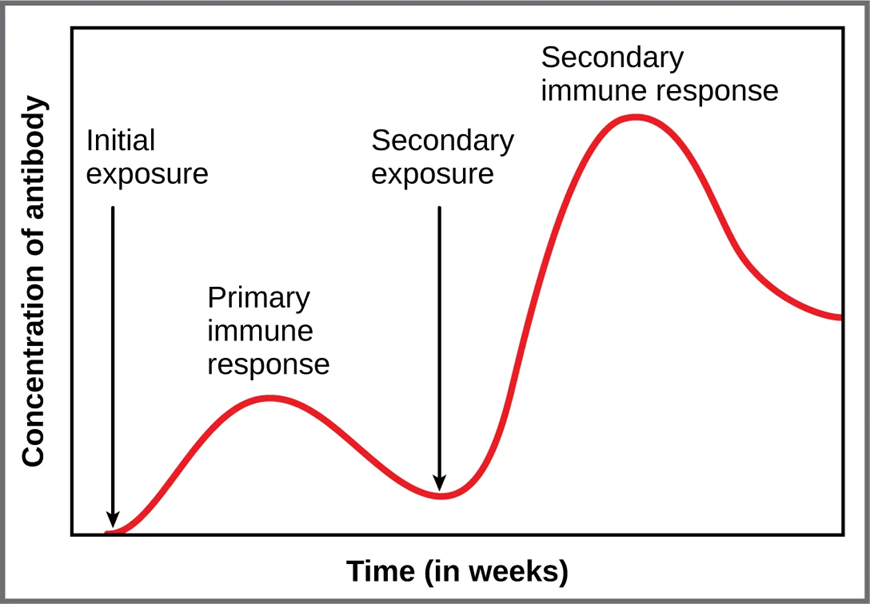
Vaccination and Immunological Memory
Vaccines are designed to stimulate the production of Memory Th cells, Memory Tc cells, and Memory B cells without causing disease. Upon subsequent pathogen exposure, these cells mount a faster, more effective response than the primary response – providing lasting protection. Memory cell pools are maintained by balanced proliferation and apoptosis, preserving antigen memory for 10 years or more.
Media Attributions
- Antibodies © Mary Ann Clark, Matthew Douglas, Jung Choi is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- APC © Mary Ann Clark, Matthew Douglas, Jung Choi is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- APC MHC1 MHC2 © . Gordon Betts, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble, Peter DeSaix is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- B Cell Activation © Fred the Oyster is licensed under a Public Domain license
- Sensitized B Cell is Activated by Helper T cell © J. Gordon Betts, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble, Peter DeSaix is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Private: 05e3c5fcb2a46e4b7b2ba915fd41360e5d69eb22 © OpenStax College is licensed under a CC BY (Attribution) license
- Primary and Secondary Immune Response Antibody Titre © J. Gordon Betts, Kelly A. Young, James A. Wise, Eddie Johnson, Brandon Poe, Dean H. Kruse, Oksana Korol, Jody E. Johnson, Mark Womble, Peter DeSaix is licensed under a CC BY-NC-SA (Attribution NonCommercial ShareAlike) license
- Poliodrops © USAID is licensed under a Public Domain license

{kind=link}
{kind=link}