Neuromuscular Control of Motion
Somatic Nervous System
J. Gordon Betts; James A. Wise; Kelly A. Young; Eddie Johnson; Dean H. Kruse; Oksana Korol; Jody E. Johnson; Mark Womble; Peter DeSaix; and Brandon Poe
Learning Objectives
By the end of this section, you will be able to:
- Describe the components of the somatic nervous system.
- Describe the different types and classifications of sensory receptors.
- Name the modalities and submodalities of the sensory systems.
- Distinguish between general and special senses.
- Describe regions of the central nervous system that contribute to somatic functions.
- Explain the stimulus-response motor pathway.
The somatic nervous system is traditionally considered a division within the peripheral nervous system. However, this misses an important point: somatic refers to a functional division, whereas peripheral refers to an anatomic division. The somatic nervous system is responsible for our conscious sensation and perception of the environment and for our responses to that perception by means of voluntary movement of skeletal muscles. Additionally, the somatic nervous system also controls some involuntary muscle movements known as reflexes.
In this chapter, we will focus on the sensory aspects of the somatic nervous system, specifically the somatosensory system. For now, just keep in mind that the somatic nervous system includes both sensation and motor responses.
Sensation
The sensory systems throughout our body share a single purpose: to help us detect important stimuli in our external environment. Stimuli from varying sources, and of different types, are detected and converted into the electrochemical signals of the nervous system. This occurs when a stimulus changes the cell membrane potential of a sensory neuron. The stimulus causes the sensory cell to produce an action potential that is relayed into the central nervous system (CNS), where it is integrated with other sensory information—or sometimes higher cognitive functions—to become a conscious perception of that stimulus. The central integration may then lead to a motor response.
An important distinction to make here is between sensation and perception, which are often mistakenly used interchangeably in everyday language. Sensation is the activation of sensory receptor cells at the level of the stimulus, for example thermoreceptors in the skin detecting heat from your hand touching a hot stove. Perception is the central processing of sensory stimuli into a meaningful pattern, for example perceiving that contact with the stove as being hot and painful. Perception is dependent on sensation, but not all sensations are perceived.
Sensory Receptors
Stimuli in the environment activate specialized receptor cells in the peripheral nervous system, for example heat activating thermoreceptors in the skin as explained above. Different types of stimuli are sensed by different types of receptor cells. Receptor cells can be classified into types on the basis of three different criteria: cell type, position, and function. Receptors can be classified structurally on the basis of cell type and their position in relation to stimuli they sense. They can also be classified functionally on the basis of the transduction of stimuli, or how the mechanical stimulus, light, or chemical changes the cell membrane potential.
Structural Receptor Classifications
The cells that interpret information about the environment can be either (1) a neuron that has a free nerve ending, with dendrites embedded in tissue that would receive a sensation; (2) a neuron that has an encapsulated ending in which the sensory nerve endings are encapsulated in connective tissue that enhances their sensitivity; or (3) a specialized receptor cell, which has distinct structural components that interpret a specific type of stimulus (Receptor Classification by Cell Type). The pain and temperature receptors in the dermis of the skin are examples of neurons that have free nerve endings. Also located in the dermis of the skin are lamellated corpuscles, neurons with encapsulated nerve endings that respond to pressure and touch. The cells in the retina that respond to light stimuli are an example of a specialized receptor, a photoreceptor.
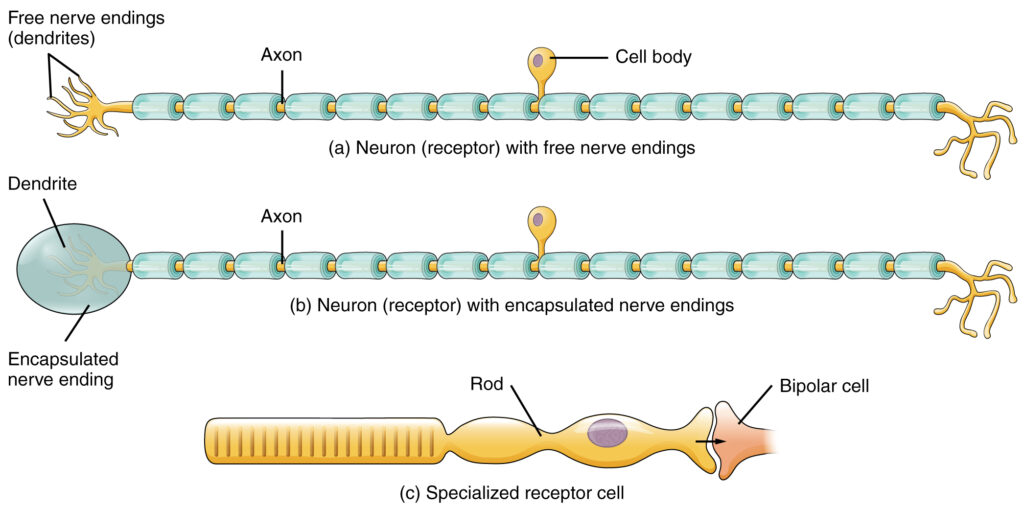
Another way that receptors can be classified is based on their location relative to the stimuli. An exteroceptor is a receptor that is located near a stimulus in the external environment, such as the somatosensory receptors that are located in the skin. An interoceptor is one that interprets stimuli from internal organs and tissues, such as the receptors that sense the increase in blood pressure in the aorta or carotid sinus. Finally, a proprioceptor is a receptor located near a moving part of the body, such as a muscle, that interprets the positions of the tissues as they move.
Functional Receptor Classifications
The third classification of receptors is by how the receptor transduces stimuli into membrane potential changes. Stimuli are of three general types. Some stimuli are ions and macromolecules that affect transmembrane receptor proteins when these chemicals diffuse across the cell membrane. Some stimuli are physical variations in the environment that affect receptor cell membrane potentials. Other stimuli include electromagnetic radiation from visible light. For humans, the only electromagnetic energy that is perceived by our eyes is visible light. Some other organisms have receptors that humans lack, such as the heat sensors of snakes, the ultraviolet light sensors of bees, or magnetic receptors in migratory birds.
Receptor cells can be further categorized on the basis of the type of stimuli they transduce. Chemical stimuli can be interpreted by a chemoreceptor that interprets chemical stimuli, such as an object’s taste or smell. Osmoreceptors respond to solute concentrations of body fluids. Additionally, pain is primarily a chemical sense that interprets the presence of chemicals from tissue damage, or similar intense stimuli, through a nociceptor. Physical stimuli, such as pressure and vibration, as well as the sensation of sound and body position (balance), are interpreted through a mechanoreceptor. Another physical stimulus that has its own type of receptor is temperature, which is sensed through a thermoreceptor that is either sensitive to temperatures above (heat) or below (cold) normal body temperature.
Sensory Modalities
Ask anyone what the senses are, and they are likely to list the five major senses—taste, smell, touch, hearing, and sight. However, these are not all of the senses. The most obvious omission from this list is balance. Also, what is referred to simply as touch can be further subdivided into pressure, vibration, stretch, and hair-follicle position, on the basis of the type of mechanoreceptors that perceive these touch sensations. Other overlooked senses include temperature perception by thermoreceptors and pain perception by nociceptors.
Within the realm of physiology, senses can be classified as either general or specific. A general sense is one that is distributed throughout the body and has receptor cells within the structures of other organs. Mechanoreceptors in the skin, muscles, or the walls of blood vessels are examples of this type. General senses often contribute to the sense of touch, as described above, or to proprioception (body movement) and kinesthesia (body movement), or to a visceral sense, which is most important to autonomic functions. A special sense is one that has a specific organ devoted to it, namely the eye, inner ear, tongue, or nose.
Each of the senses is referred to as a sensory modality. Modality refers to the way that information is encoded, which is similar to the idea of transduction. The main sensory modalities can be described on the basis of how each is transduced. The chemical senses are taste and smell. The general sense that is usually referred to as touch includes chemical sensation in the form of nociception, or pain. Pressure, vibration, muscle stretch, and the movement of hair by an external stimulus, are all sensed by mechanoreceptors. Hearing and balance are also sensed by mechanoreceptors. Finally, vision involves the activation of photoreceptors.
Listing all the different sensory modalities, which can number as many as 17, involves separating the five major senses into more specific categories, or submodalities, of the larger sense. An individual sensory modality represents the sensation of a specific type of stimulus. For example, the general sense of touch, which is known as somatosensation, can be separated into light pressure, deep pressure, vibration, itch, pain, temperature, or hair movement.
Somatic Nervous System and the CNS
Specific regions of the CNS coordinate different somatic processes using sensory inputs and motor outputs of peripheral nerves. A simple case is the withdrawal reflex caused by a synapse between a dorsal sensory neuron axon and a motor neuron in the ventral horn like we discussed before. More complex arrangements are possible to integrate peripheral sensory information with higher processes. The important regions of the CNS that play a role in somatic processes can be separated into the spinal cord brain stem, diencephalon, cerebral cortex, and subcortical structures.
A sensory pathway that carries peripheral sensations to the brain is referred to as an ascending pathway, or ascending tract. The various sensory modalities each follow specific pathways through the CNS. Tactile and other somatosensory stimuli activate receptors in the skin, muscles, tendons, and joints throughout the entire body. However, the somatosensory pathways are divided into two separate systems on the basis of the location of the receptor neurons. Somatosensory stimuli from below the neck pass along the sensory pathways of the spinal cord, whereas somatosensory stimuli from the head and neck travel through the cranial nerves—specifically, the trigeminal system.
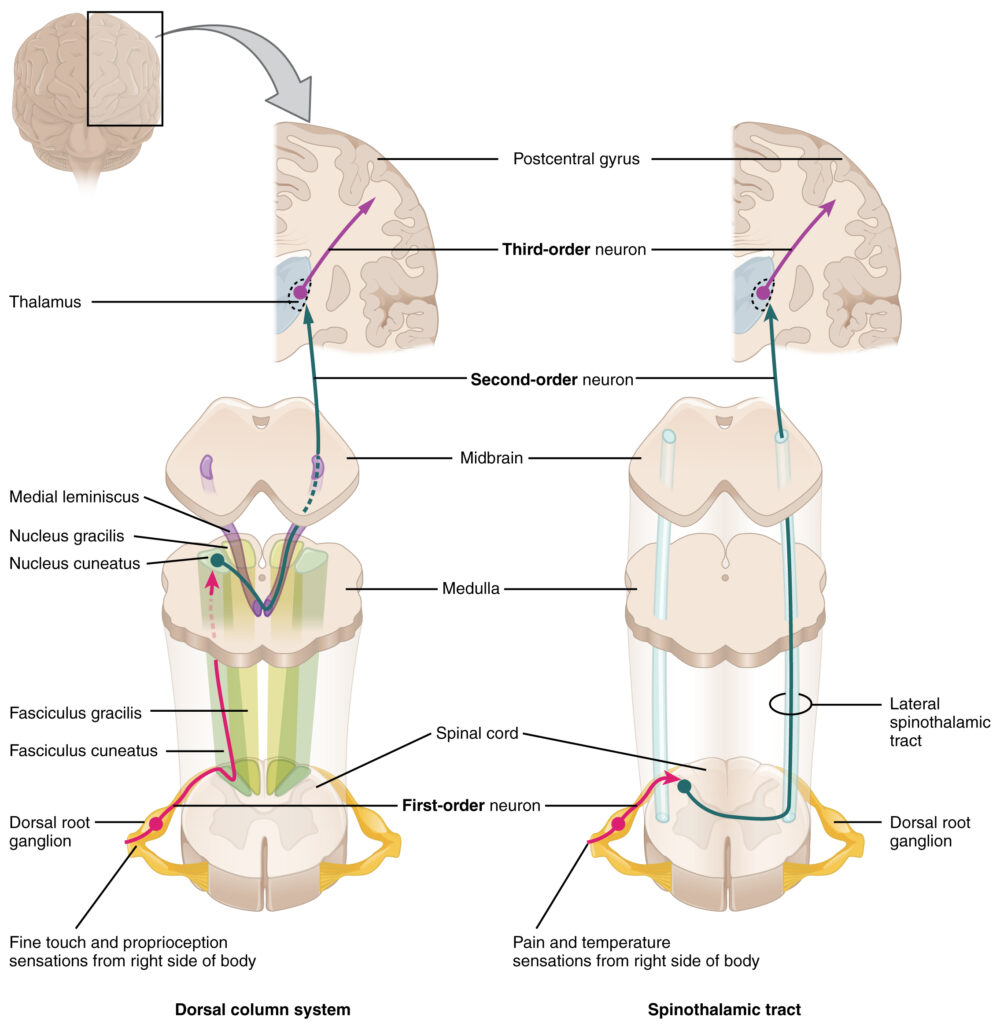
The dorsal column system (sometimes referred to as the dorsal column–medial lemniscus) and the spinothalamic tract are two major pathways that bring sensory information to the brain. The sensory pathways in each of these systems are composed of three successive neurons.
The dorsal column system begins with the axon of a dorsal root ganglion neuron entering the dorsal root and joining the dorsal column white matter in the spinal cord. As axons of this pathway enter the dorsal column, they take on a positional arrangement so that axons from lower levels of the body position themselves medially, whereas axons from upper levels of the body position themselves laterally. The dorsal column is separated into two component tracts, the fasciculus gracilis that contains axons from the legs and lower body, and the fasciculus cuneatus that contains axons from the upper body and arms.
The axons in the dorsal column terminate in the nuclei of the medulla, where each synapses with the second neuron in their respective pathway. The nucleus gracilis is the target of fibers in the fasciculus gracilis, whereas the nucleus cuneatus is the target of fibers in the fasciculus cuneatus. The second neuron in the system projects from one of the two nuclei and then decussates, or crosses the midline of the medulla. These axons then continue to ascend the brain stem on the contralateral (opposite) side as a bundle called the medial lemniscus. These axons terminate in the thalamus, where each synapses with the third neuron in their respective pathway. The third neuron in the system projects its axons to the postcentral gyrus of the cerebral cortex, where somatosensory stimuli are initially processed and the conscious perception of the stimulus occurs.
The other major pathway, the spinothalamic tract, also begins with neurons in a dorsal root ganglion. These neurons extend their axons to the dorsal horn, where they synapse with the second neuron in their respective pathway. The name “spinothalamic” comes from this second neuron, which has its cell body in the spinal cord gray matter and connects to the thalamus. Axons from these second neurons then decussate within the spinal cord and ascend to the brain and enter the thalamus, where each synapses with the third neuron in its respective pathway. The neurons in the thalamus then project their axons to the spinothalamic tract, which synapses in the postcentral gyrus of the cerebral cortex.
These two systems are similar in that they both begin with dorsal root ganglion cells, as with most general sensory information. The dorsal column system is primarily responsible for touch sensations and proprioception, whereas the spinothalamic tract pathway is primarily responsible for pain and temperature sensations. Another similarity is that the second neurons in both of these pathways are contralateral because they project across the midline to the other side of the brain or spinal cord. In the dorsal column system, this decussation takes place in the brain stem; in the spinothalamic pathway, it takes place in the spinal cord at the same spinal cord level at which the information entered. The third neurons in the two pathways are essentially the same. In both, the second neuron synapses in the thalamus, and the thalamic neuron projects to the somatosensory cortex.
The trigeminal pathway carries somatosensory information from the face, head, mouth, and nasal cavity. As with the previously discussed nerve tracts, the sensory pathways of the trigeminal pathway each involve three successive neurons. First, axons from the trigeminal ganglion enter the brain stem at the level of the pons. These axons project to one of three locations. The spinal trigeminal nucleus of the medulla receives information similar to that carried by spinothalamic tract, such as pain and temperature sensations. Other axons go to either the chief sensory nucleus in the pons or the mesencephalic nuclei in the midbrain. These nuclei receive information like that carried by the dorsal column system, such as touch, pressure, vibration, and proprioception. Axons from the second neuron decussate and ascend to the thalamus along the trigeminothalamic tract. In the thalamus, each axon synapses with the third neuron in its respective pathway. Axons from the third neuron then project from the thalamus to the primary somatosensory cortex of the cerebrum.
Diencephalon
The diencephalon is beneath the cerebrum and includes the thalamus and hypothalamus. In the somatic nervous system, the thalamus is an important relay for communication between the cerebrum and the rest of the nervous system. The hypothalamus has both somatic and autonomic functions. In addition, the hypothalamus communicates with the limbic system, which controls emotions and memory functions.
Sensory input to the thalamus comes from most of the special senses and ascending somatosensory tracts. Each sensory system is relayed through a particular nucleus in the thalamus. The thalamus is a required transfer point for most sensory tracts that reach the cerebral cortex, where conscious sensory perception begins. The one exception to this rule is the olfactory system. The olfactory tract axons from the olfactory bulb project directly to the cerebral cortex, along with the limbic system and hypothalamus.
Cortical Processing
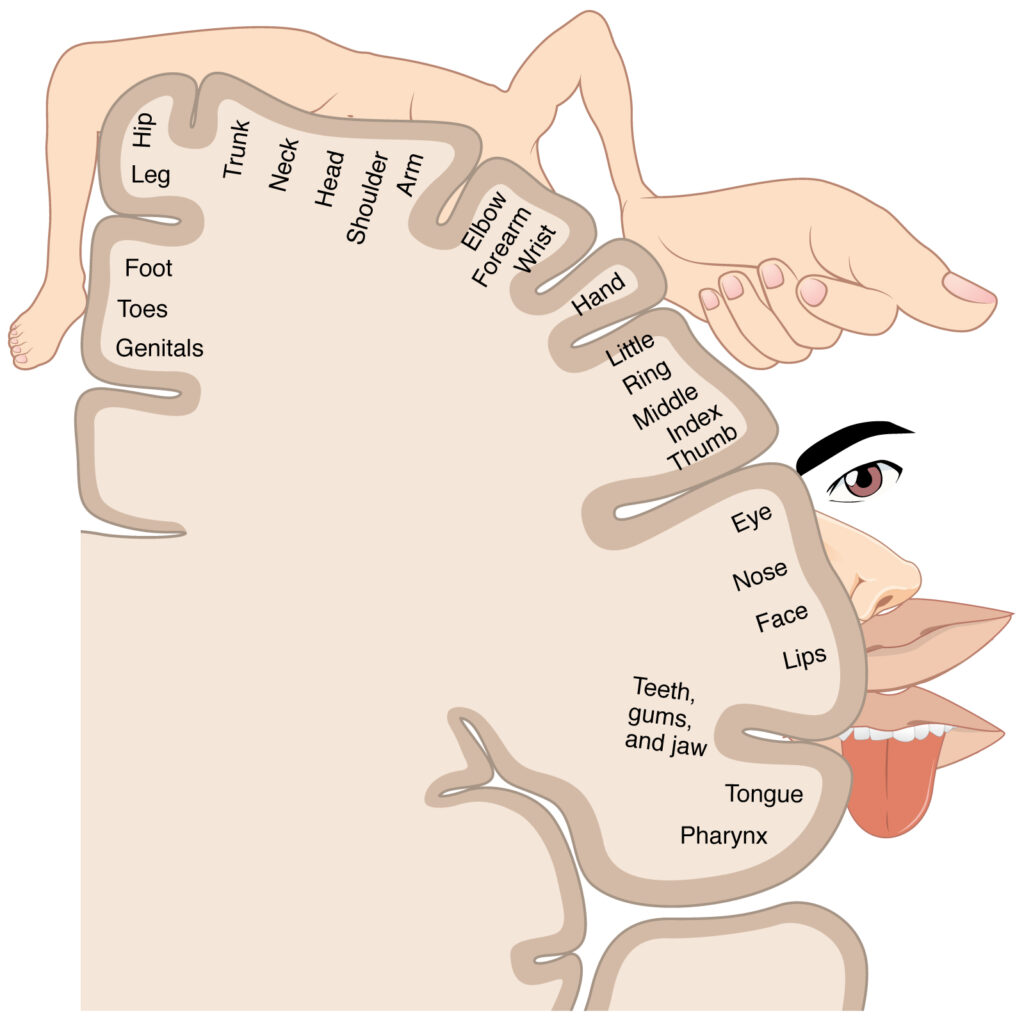
As described earlier, many of the sensory axons are positioned in the same way as their corresponding receptor cells in the body. This allows identification of the position of a stimulus on the basis of which receptor cells are sending information. The cerebral cortex also maintains this sensory topography in the particular areas of the cortex that correspond to the position of the receptor cells. The somatosensory cortex provides an example in which, in essence, the locations of the somatosensory receptors in the body are mapped onto the somatosensory cortex. This mapping is often depicted using a sensory homunculus (The Sensory Homunculus).
The term homunculus comes from the Latin word for “little man” and refers to a map of the human body that is laid across a portion of the cerebral cortex. In the somatosensory cortex, the external genitals, feet, and lower legs are represented on the medial face of the gyrus within the longitudinal fissure. As the gyrus curves out of the fissure and along the surface of the parietal lobe, the body map continues through the thighs, hips, trunk, shoulders, arms, and hands. The head and face are just lateral to the fingers as the gyrus approaches the lateral sulcus. The representation of the body in this topographical map is medial to lateral from the lower to the upper body. Note that this correspondence does not result in a perfectly miniature scale version of the body, but rather exaggerates the more sensitive areas of the body, such as the fingers and lower face. Less sensitive areas of the body, such as the shoulders and back, are mapped to smaller areas on the cortex.
Adaption
This chapter was adapted by Valerie Swanston from the following text:
Sensory Perception in Anatomy and Physiology by OSCRiceUniversity is licensed under a Creative Commons Attribution 4.0 International License
Media Attributions
- 1401_Receptor_Types-1 © OSCRiceUniversity is licensed under a CC BY (Attribution) license
- 1417_Ascending_Pathways_of_Spinal_Cord © OSCRiceUniversity is licensed under a CC BY (Attribution) license
- 1421_Sensory_Homunculus © OSCRiceUniversity is licensed under a CC BY (Attribution) license
